

Multiregion neuronal activity: the forest and the trees
- PMID: 36192596
- PMCID: PMC10327445
- DOI: 10.1038/s41583-022-00634-0
Multiregion neuronal activity: the forest and the trees
Abstract
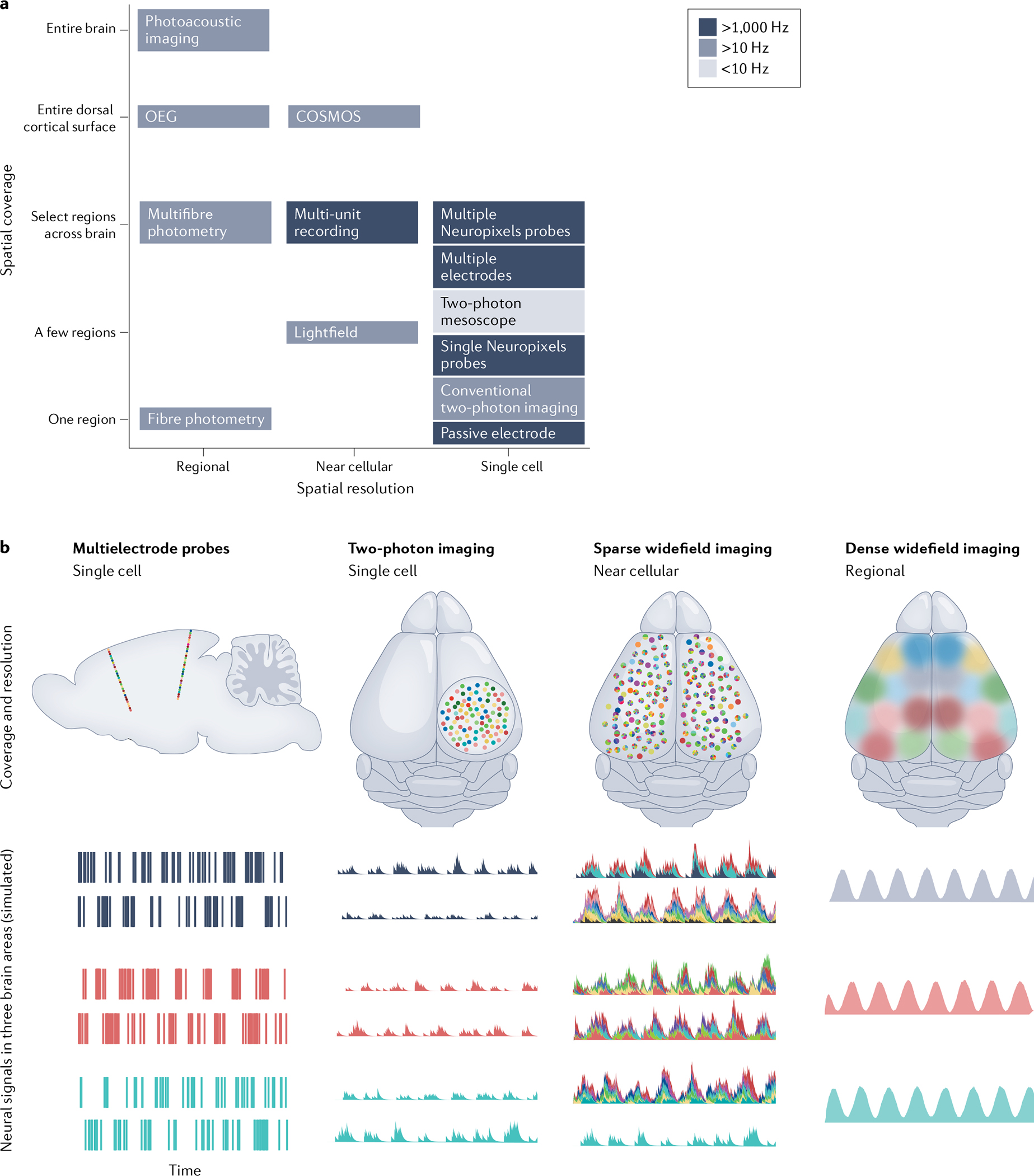
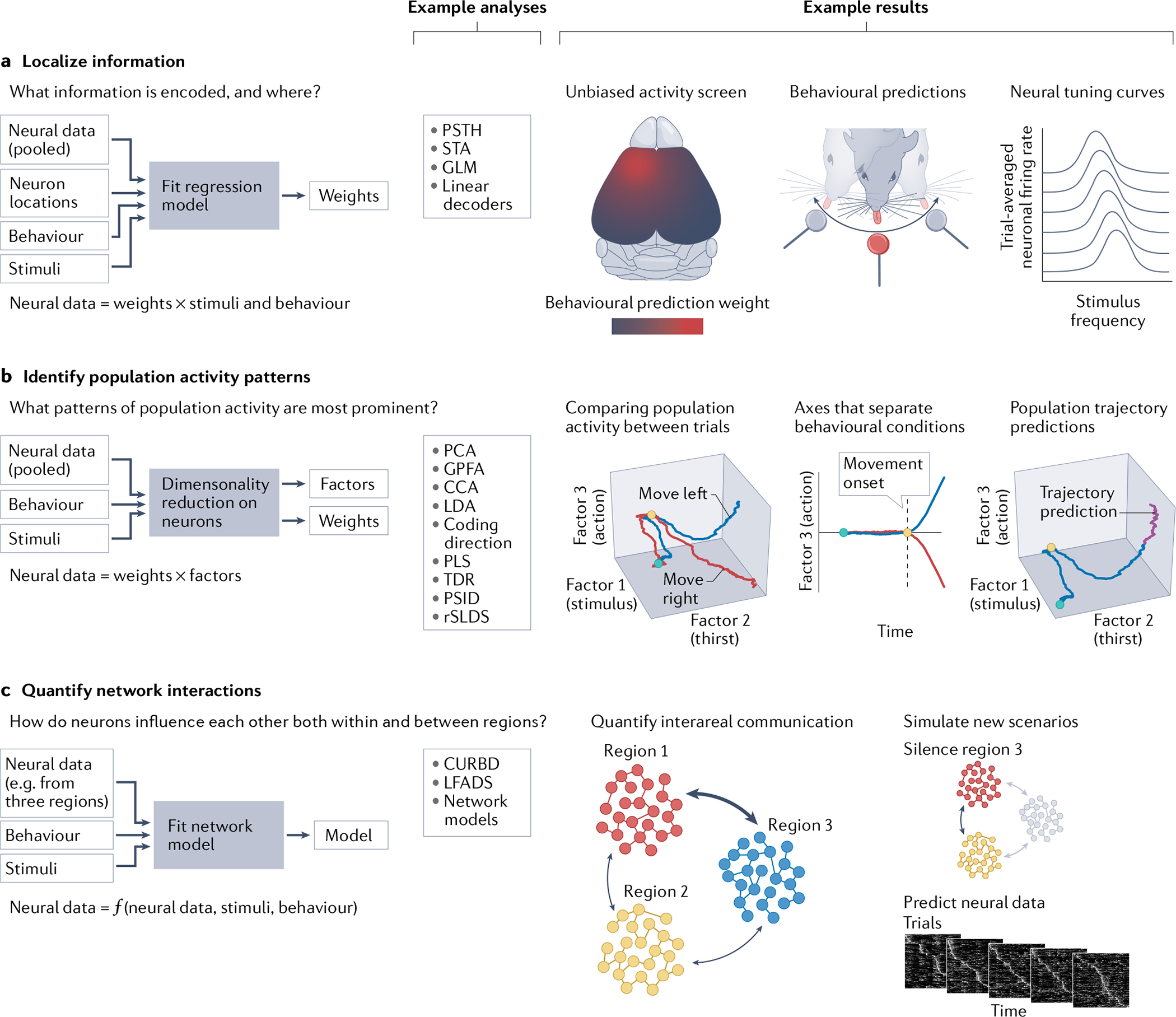
The past decade has witnessed remarkable advances in the simultaneous measurement of neuronal activity across many brain regions, enabling fundamentally new explorations of the brain-spanning cellular dynamics that underlie sensation, cognition and action. These recently developed multiregion recording techniques have provided many experimental opportunities, but thoughtful consideration of methodological trade-offs is necessary, especially regarding field of view, temporal acquisition rate and ability to guarantee cellular resolution. When applied in concert with modern optogenetic and computational tools, multiregion recording has already made possible fundamental biological discoveries - in part via the unprecedented ability to perform unbiased neural activity screens for principles of brain function, spanning dozens of brain areas and from local to global scales.
© 2022. Springer Nature Limited.
Conflict of interest statement
Competing interests
The authors declare no competing financial interests
Figures

References
-
- The Event Horizon Telescope Collaboration. et al. First M87 event horizon telescope results. I. The shadow of the supermassive black hole. Astrophys. J. Lett. 875, L1 (2019).
-
- Galilei G Sidereus Nuncius (Univ. Chicago Press, 1610).
-
- Adrian ED The Basis of Sensation (WW Norton & Co, 1928).
-
- Woodbury JW & Patton HD in Cold Spring Harbor Symposia on Quantitative Biology vol. 17, 185–188 (Cold Spring Harbor Laboratory Press, 1952). - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
