

Life cycle and functional genomics of the unicellular red alga Galdieria for elucidating algal and plant evolution and industrial use
- PMID: 36194630
- PMCID: PMC9565259
- DOI: 10.1073/pnas.2210665119
Life cycle and functional genomics of the unicellular red alga Galdieria for elucidating algal and plant evolution and industrial use
Abstract
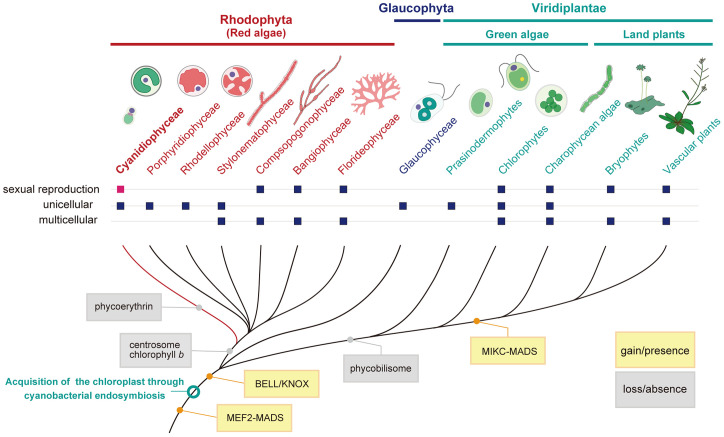
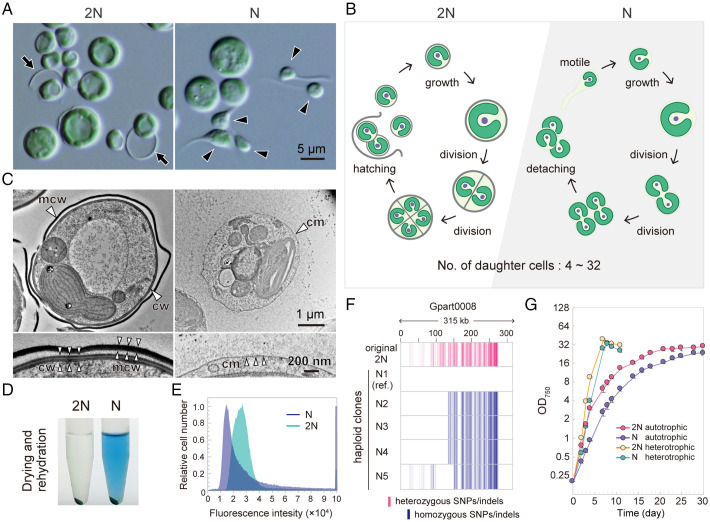
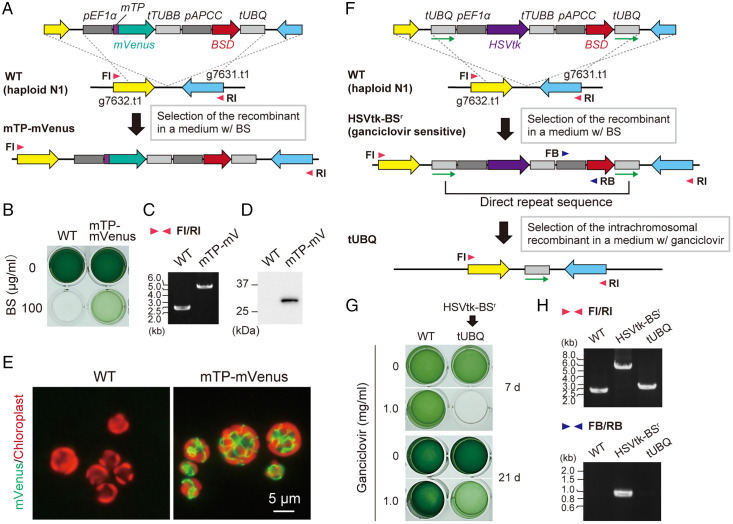
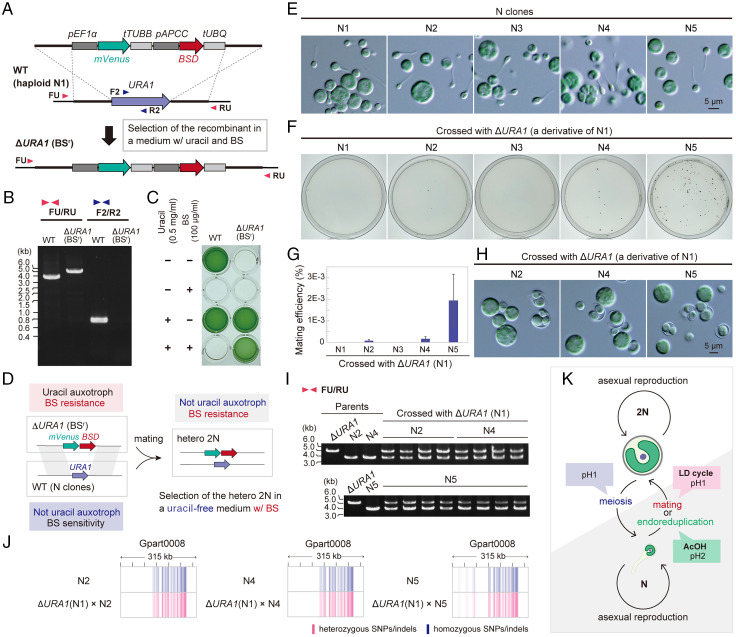
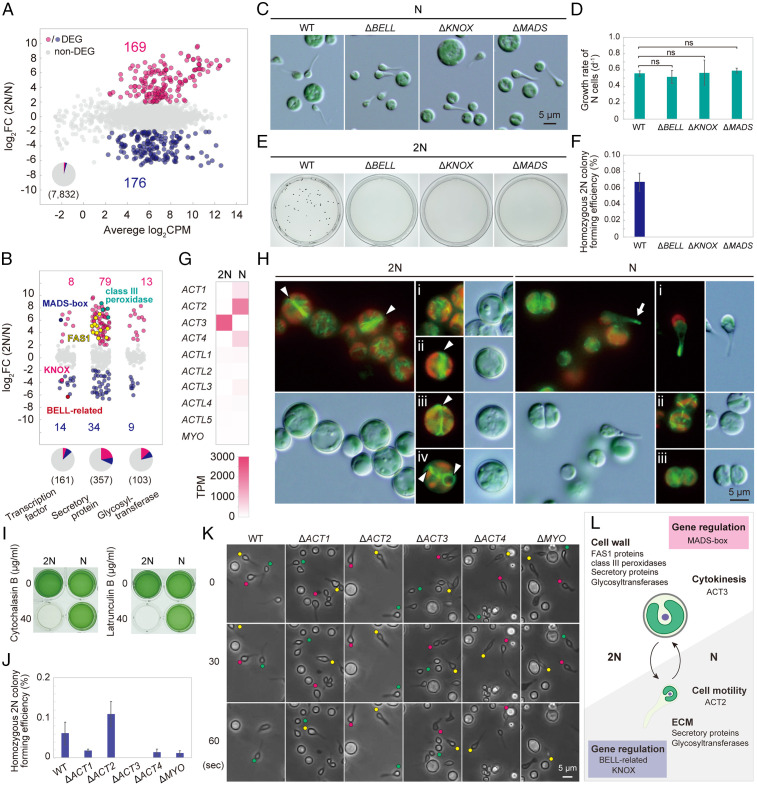
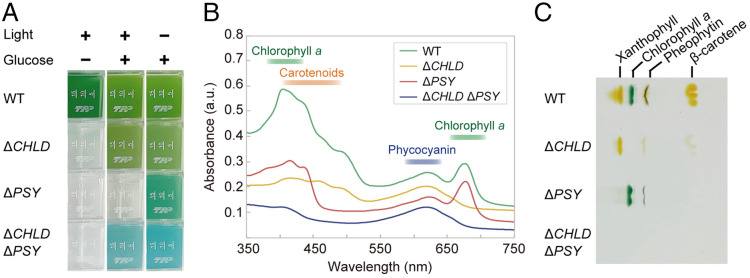
Sexual reproduction is widespread in eukaryotes; however, only asexual reproduction has been observed in unicellular red algae, including Galdieria, which branched early in Archaeplastida. Galdieria possesses a small genome; it is polyextremophile, grows either photoautotrophically, mixotrophically, or heterotrophically, and is being developed as an industrial source of vitamins and pigments because of its high biomass productivity. Here, we show that Galdieria exhibits a sexual life cycle, alternating between cell-walled diploid and cell wall-less haploid, and that both phases can proliferate asexually. The haploid can move over surfaces and undergo self-diploidization or generate heterozygous diploids through mating. Further, we prepared the whole genome and a comparative transcriptome dataset between the diploid and haploid and developed genetic tools for the stable gene expression, gene disruption, and selectable marker recycling system using the cell wall-less haploid. The BELL/KNOX and MADS-box transcription factors, which function in haploid-to-diploid transition and development in plants, are specifically expressed in the haploid and diploid, respectively, and are involved in the haploid-to-diploid transition in Galdieria, providing information on the missing link of the sexual life cycle evolution in Archaeplastida. Four actin genes are differently involved in motility of the haploid and cytokinesis in the diploid, both of which are myosin independent and likely reflect ancestral roles of actin. We have also generated photosynthesis-deficient mutants, such as blue-colored cells, which were depleted in chlorophyll and carotenoids, for industrial pigment production. These features of Galdieria facilitate the understanding of the evolution of algae and plants and the industrial use of microalgae.
Keywords: Galdieria; actin; homeobox; microalgae; sexual reproduction.
Conflict of interest statement
Competing interest statement: The Japan Science and Technology Agency has filed patent applications related to the generation and maintenance of
Figures
Comment in
-
Discovery of sex in an extremophilic red alga.Proc Natl Acad Sci U S A. 2022 Nov;119(44):e2216012119. doi: 10.1073/pnas.2216012119. Epub 2022 Oct 21. Proc Natl Acad Sci U S A. 2022. PMID: 36269868 Free PMC article. No abstract available.
Similar articles
-
Maintenance of Complex Life Cycles Via Cryptic Differences In The Ecophysiology Of Haploid And Diploid Spores Of An Isomorphic Red Alga1.J Phycol. 2020 Feb;56(1):159-169. doi: 10.1111/jpy.12930. Epub 2019 Nov 20. J Phycol. 2020. PMID: 31595519
-
Comparative genomics of two closely related unicellular thermo-acidophilic red algae, Galdieria sulphuraria and Cyanidioschyzon merolae, reveals the molecular basis of the metabolic flexibility of Galdieria sulphuraria and significant differences in carbohydrate metabolism of both algae.Plant Physiol. 2005 Feb;137(2):460-74. doi: 10.1104/pp.104.051169. Plant Physiol. 2005. PMID: 15710685 Free PMC article.
-
Evolution in the Cycles of Life.Annu Rev Genet. 2016 Nov 23;50:133-154. doi: 10.1146/annurev-genet-120215-035227. Epub 2016 Sep 8. Annu Rev Genet. 2016. PMID: 27617970 Review.
-
Transcriptome analysis of functional differentiation between haploid and diploid cells of Emiliania huxleyi, a globally significant photosynthetic calcifying cell.Genome Biol. 2009;10(10):R114. doi: 10.1186/gb-2009-10-10-r114. Epub 2009 Oct 15. Genome Biol. 2009. PMID: 19832986 Free PMC article.
-
Life cycle and reproduction dynamics of Bangiales in response to environmental stresses.Semin Cell Dev Biol. 2023 Jan 30;134:14-26. doi: 10.1016/j.semcdb.2022.04.004. Epub 2022 Apr 12. Semin Cell Dev Biol. 2023. PMID: 35428563 Review.
Cited by
-
Temporal dynamics in a red alga dominated geothermal feature in Yellowstone National Park.ISME Commun. 2024 Dec 3;4(1):ycae151. doi: 10.1093/ismeco/ycae151. eCollection 2024 Jan. ISME Commun. 2024. PMID: 39711979 Free PMC article.
-
Development of a rapamycin-inducible protein-knockdown system in the unicellular red alga Cyanidioschyzon merolae.Plant Physiol. 2024 Sep 2;196(1):77-94. doi: 10.1093/plphys/kiae316. Plant Physiol. 2024. PMID: 38833589 Free PMC article.
-
Cell size and selection for stress-induced cell fusion in unicellular eukaryotes.PLoS Comput Biol. 2025 Apr 8;21(4):e1012418. doi: 10.1371/journal.pcbi.1012418. eCollection 2025. PLoS Comput Biol. 2025. PMID: 40198726 Free PMC article.
-
Costs of photosynthesis and cellular remodeling in trophic transitions of the unicellular red alga Galdieria partita.Commun Biol. 2025 Jun 7;8(1):891. doi: 10.1038/s42003-025-08284-5. Commun Biol. 2025. PMID: 40483364 Free PMC article.
-
Development and application of haploid embryonic stem cells.Stem Cell Res Ther. 2024 Apr 23;15(1):116. doi: 10.1186/s13287-024-03727-y. Stem Cell Res Ther. 2024. PMID: 38654389 Free PMC article. Review.
References
-
- Seckbach J., Ed., Evolutionary Pathways and Enigmatic Algae: Cyanidium caldarium (Rhodophyta) and Related Cells (Springer Science+Business Media, Dordrecht, The Netherlands, 1981).
-
- Yoon H. S., Hackett J. D., Ciniglia C., Pinto G., Bhattacharya D., A molecular timeline for the origin of photosynthetic eukaryotes. Mol. Biol. Evol. 21, 809–818 (2004). - PubMed
-
- Matsuzaki M., et al. , Genome sequence of the ultrasmall unicellular red alga Cyanidioschyzon merolae 10D. Nature 428, 653–657 (2004). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
