

Macrophages play a key role in tissue repair and regeneration
- PMID: 36196399
- PMCID: PMC9527023
- DOI: 10.7717/peerj.14053
Macrophages play a key role in tissue repair and regeneration
Abstract
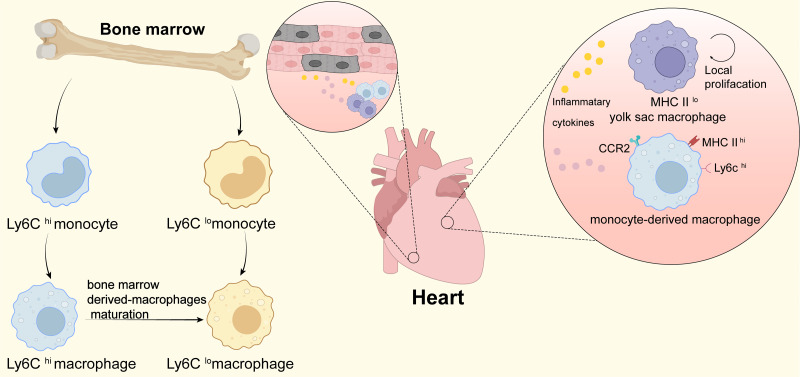
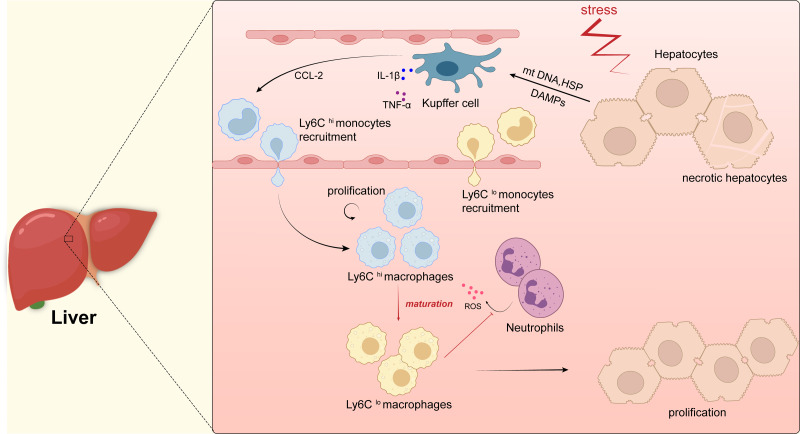
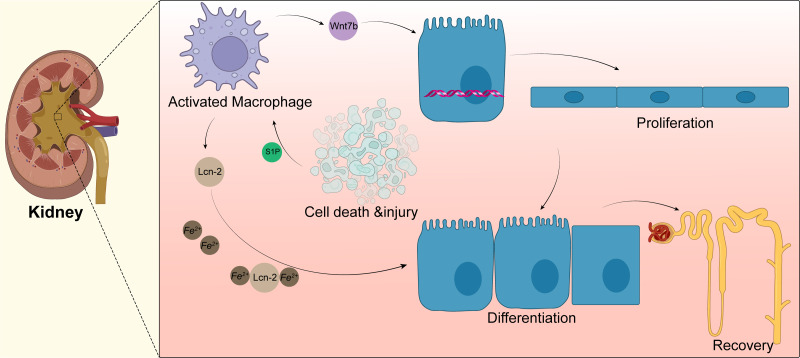
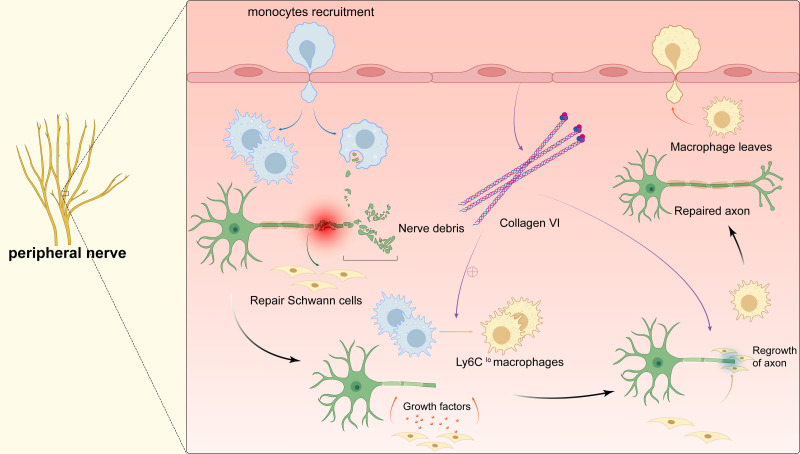
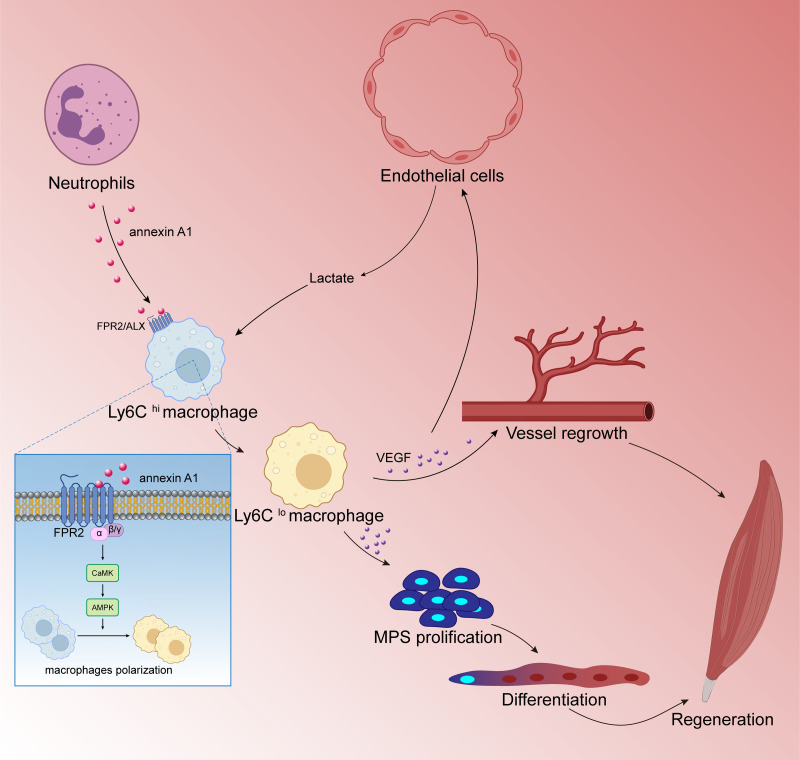
Tissue regeneration after body injury has always been a complex problem to resolve for mammals. In adult mammals, the repair process after tissue injury is often accompanied by continuous and extensive fibrosis, which leads to scars. This process has been shown to severely hinder regeneration. Macrophages, as widely distributed innate immune cells, not only play an important role in various pathological processes, but also participate in the repair process before tissue regeneration and coordinate the regeneration process after repair. This review will discuss the various forms and indispensability of macrophages involved in repair and regeneration, and how macrophages play a role in the repair and regeneration of different tissues.
Keywords: Cytokine; Polarization; Regeneration; Tissue repair; Macrophage.
©2022 Yu et al.
Conflict of interest statement
The authors declare there are no competing interests.
Figures
References
-
- Antoniades CG, Quaglia A, Taams LS, Mitry RR, Hussain M, Abeles R, Possamai LA, Bruce M, McPhail M, Starling C, Wagner B, Barnardo A, Pomplun S, Auzinger G, Bernal W, Heaton N, Vergani D, Thursz MR, Wendon J. Source and characterization of hepatic macrophages in acetaminophen-induced acute liver failure in humans. Hepatology. 2012;56:735–746. doi: 10.1002/hep.25657. - DOI - PubMed
-
- Baeck C, Wehr A, Karlmark KR, Heymann F, Vucur M, Gassler N, Huss S, Klussmann S, Eulberg D, Luedde T, Trautwein C, Tacke F. Pharmacological inhibition of the chemokine CCL2 (MCP-1) diminishes liver macrophage infiltration and steatohepatitis in chronic hepatic injury. Gut. 2012;61:416–426. doi: 10.1136/gutjnl-2011-300304. - DOI - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
