

Specialized germline P-bodies are required to specify germ cell fate in Caenorhabditis elegans embryos
- PMID: 36196602
- PMCID: PMC9686995
- DOI: 10.1242/dev.200920
Specialized germline P-bodies are required to specify germ cell fate in Caenorhabditis elegans embryos
Abstract
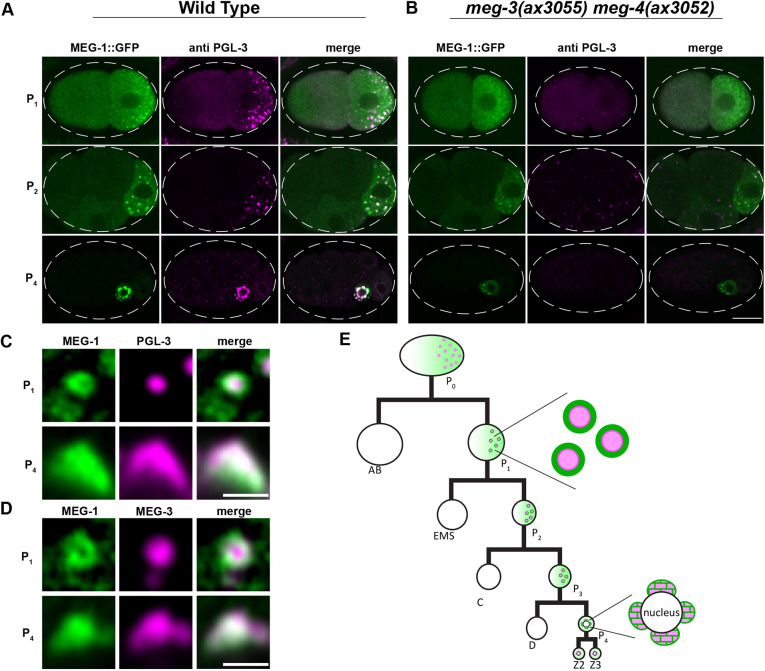
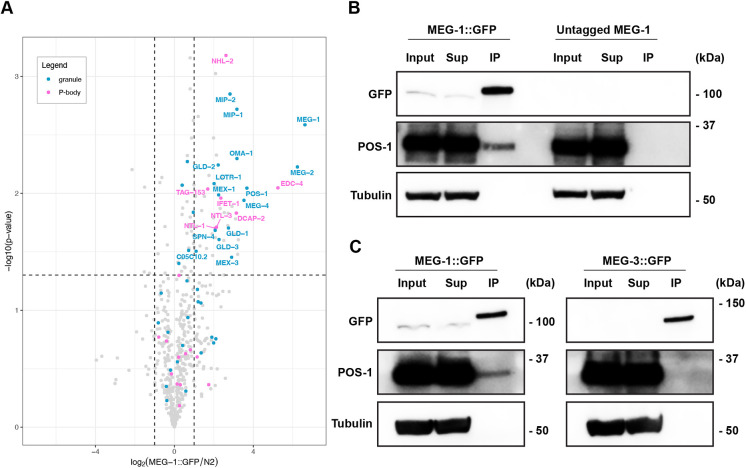
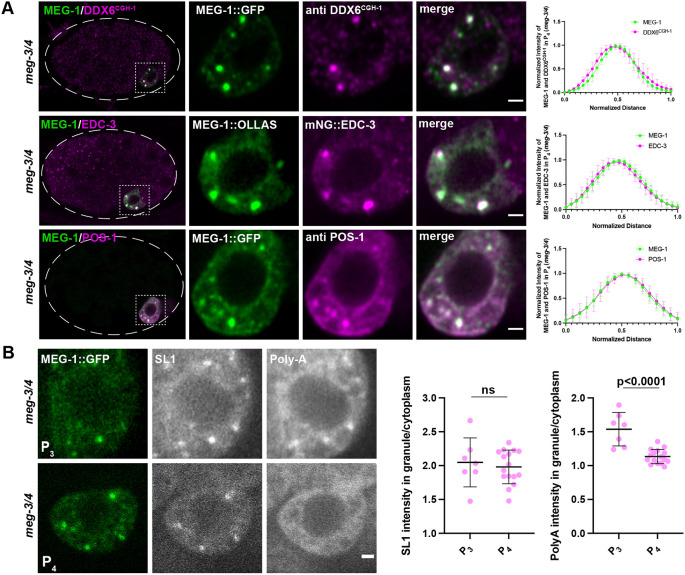
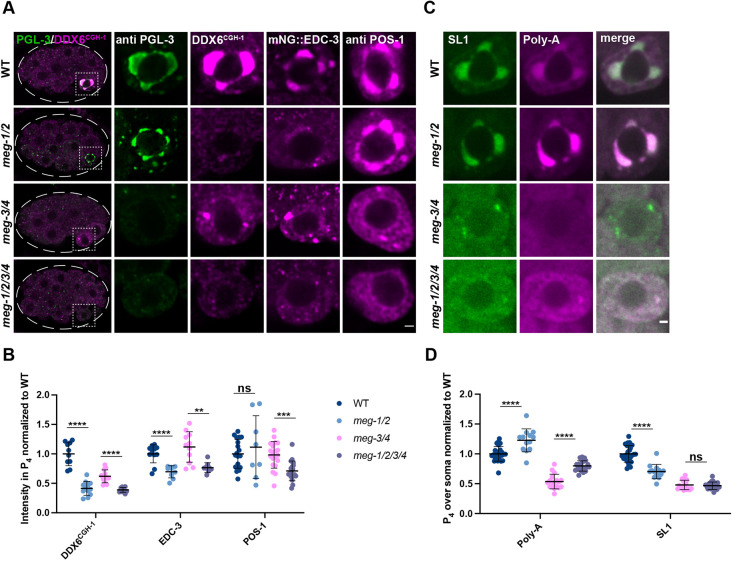
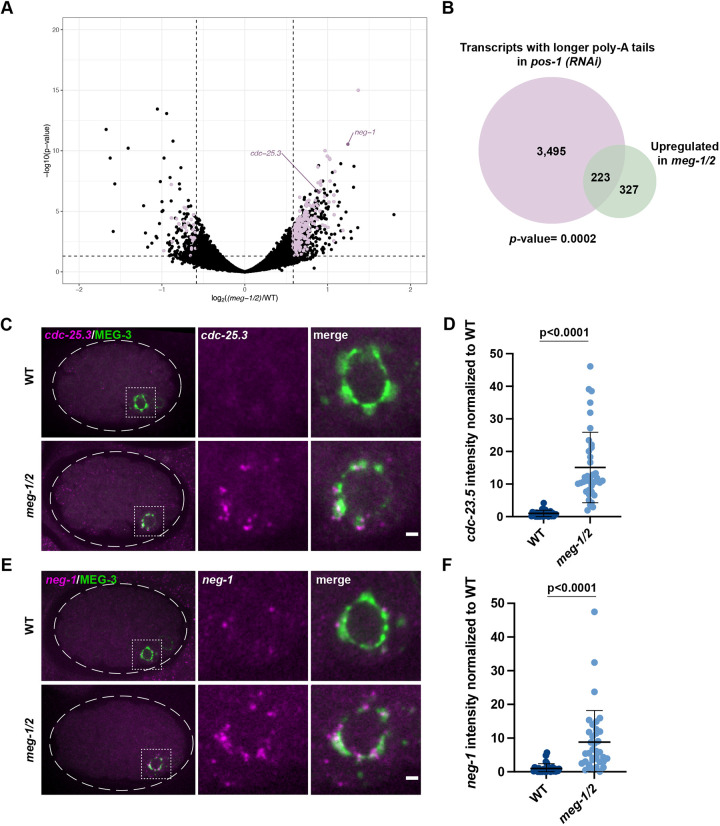
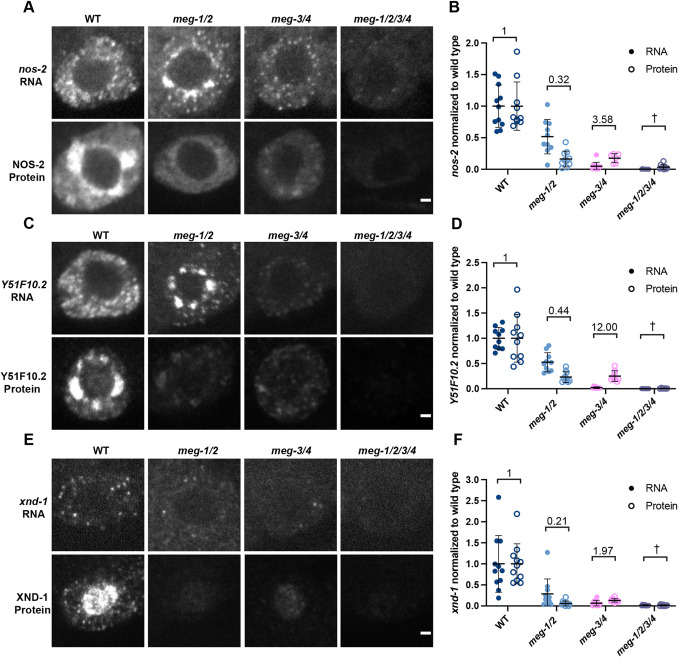
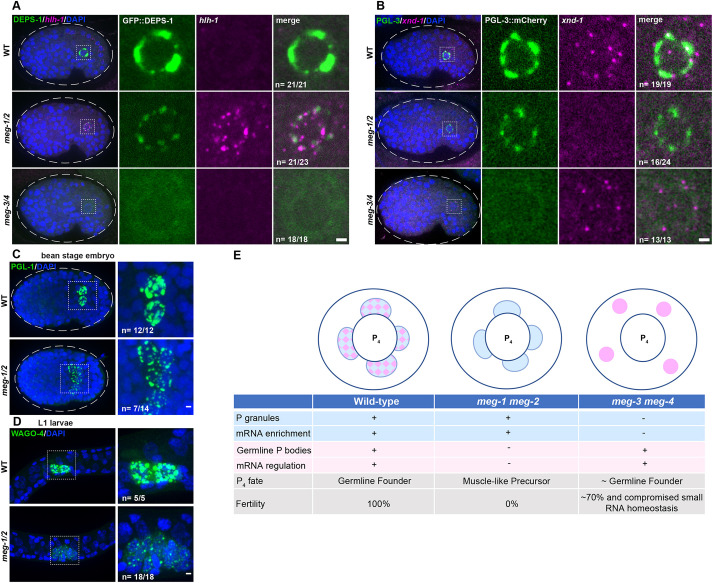
In animals with germ plasm, specification of the germline involves 'germ granules', cytoplasmic condensates that enrich maternal transcripts in the germline founder cells. In Caenorhabditis elegans embryos, P granules enrich maternal transcripts, but surprisingly P granules are not essential for germ cell fate specification. Here, we describe a second condensate in the C. elegans germ plasm. Like canonical P-bodies found in somatic cells, 'germline P-bodies' contain regulators of mRNA decapping and deadenylation and, in addition, the intrinsically-disordered proteins MEG-1 and MEG-2 and the TIS11-family RNA-binding protein POS-1. Embryos lacking meg-1 and meg-2 do not stabilize P-body components, misregulate POS-1 targets, mis-specify the germline founder cell and do not develop a germline. Our findings suggest that specification of the germ line involves at least two distinct condensates that independently enrich and regulate maternal mRNAs in the germline founder cells. This article has an associated 'The people behind the papers' interview.
Keywords: C. elegans; Germ plasm; Germline; P-bodies; Primordial germ cells; RNP granules.
© 2022. Published by The Company of Biologists Ltd.
Conflict of interest statement
Competing interests G.S. serves on the Scientific Advisory Board of Dewpoint Therapeutics. The remaining authors declare no competing interests.
Figures
References
-
- Alessi, A. F., Khivansara, V., Han, T., Freeberg, M. A., Moresco, J. J., Tu, P. G., Montoye, E., Yates, J. R., III, Karp, X. and Kim, J. K. (2015). Casein kinase II promotes target silencing by miRISC through direct phosphorylation of the DEAD-box RNA helicase CGH-1. Proc. Natl. Acad. Sci. USA 112, E7213-E7222. 10.1073/pnas.1509499112 - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
