

NOS1 mutations cause hypogonadotropic hypogonadism with sensory and cognitive deficits that can be reversed in infantile mice
- PMID: 36197968
- PMCID: PMC7613826
- DOI: 10.1126/scitranslmed.abh2369
NOS1 mutations cause hypogonadotropic hypogonadism with sensory and cognitive deficits that can be reversed in infantile mice
Abstract
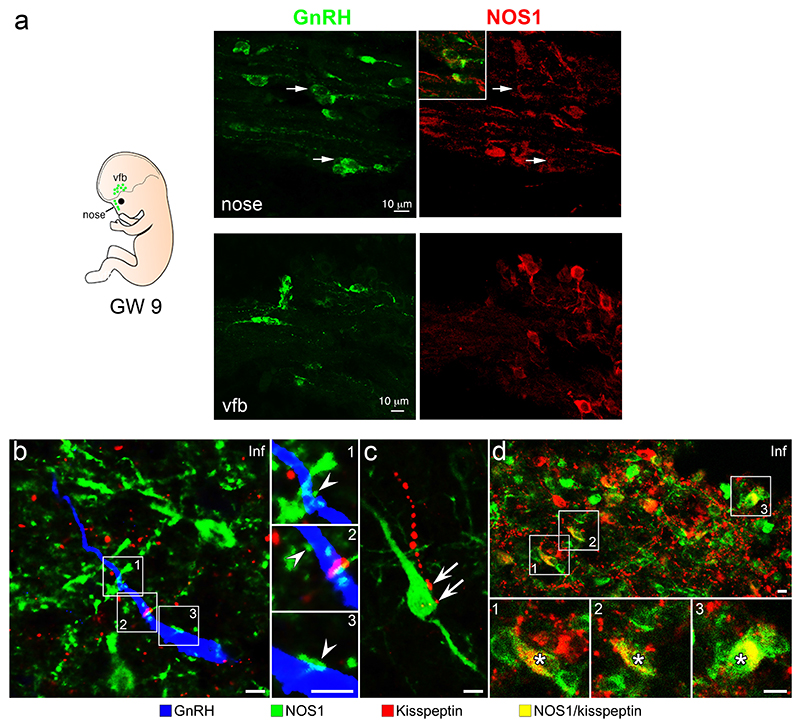
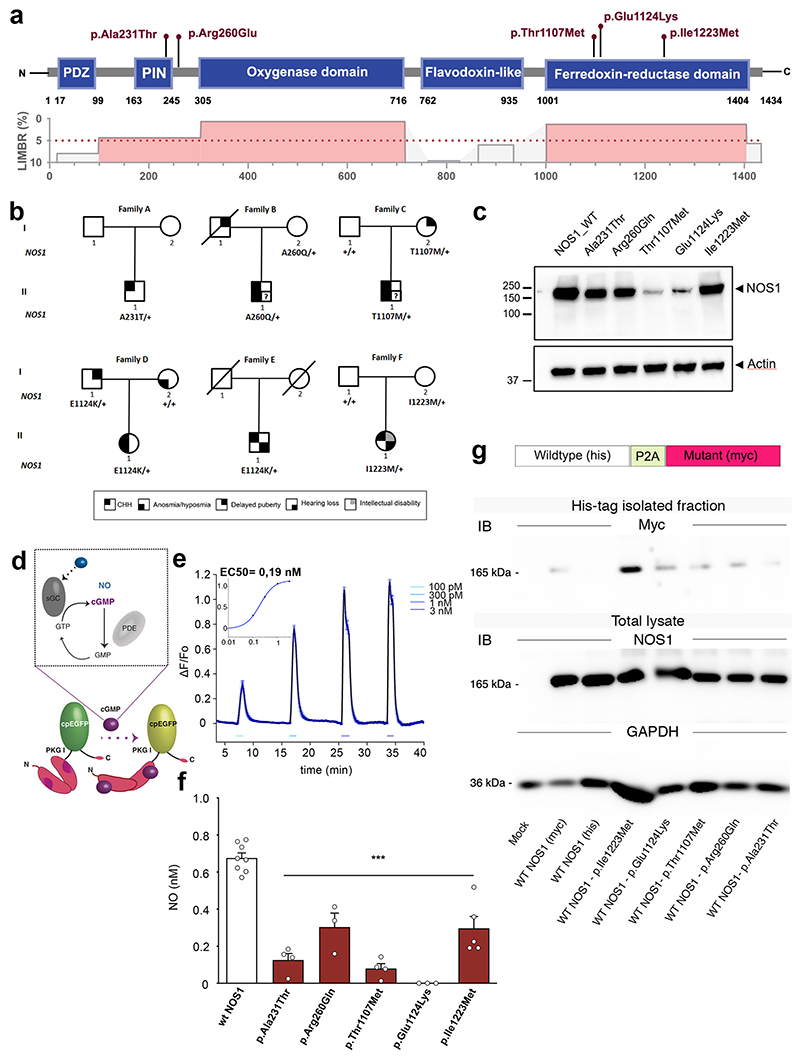
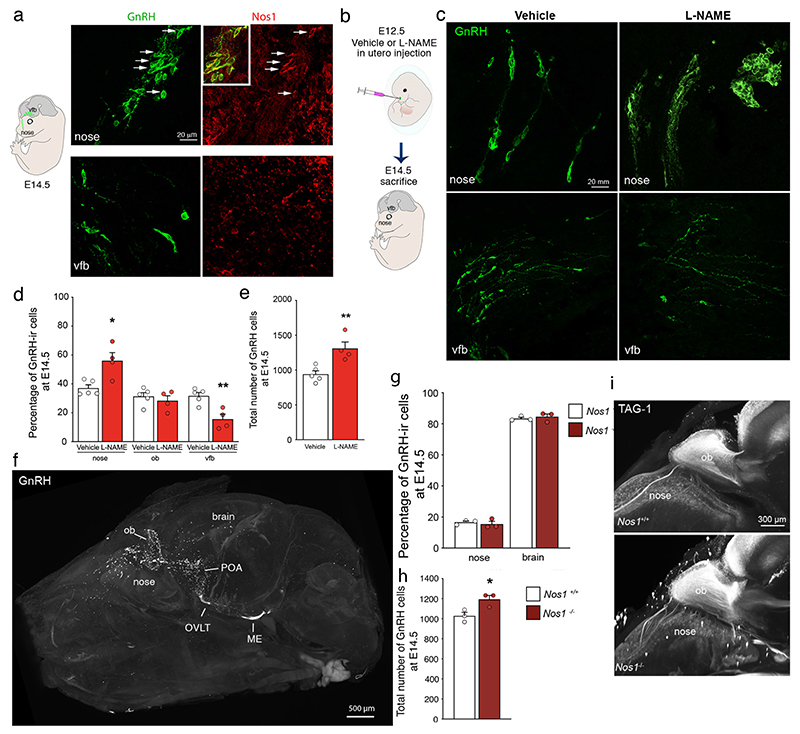
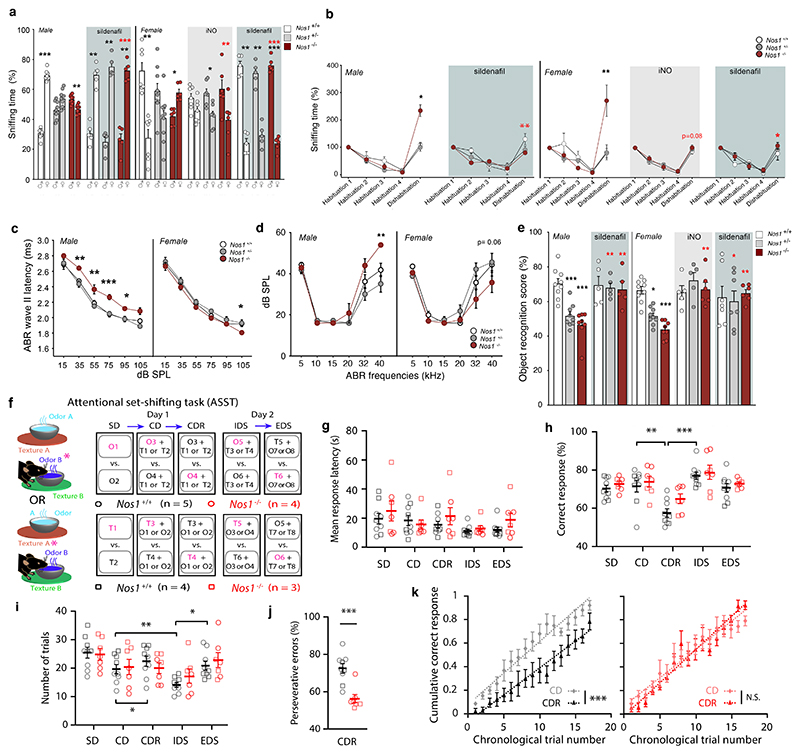
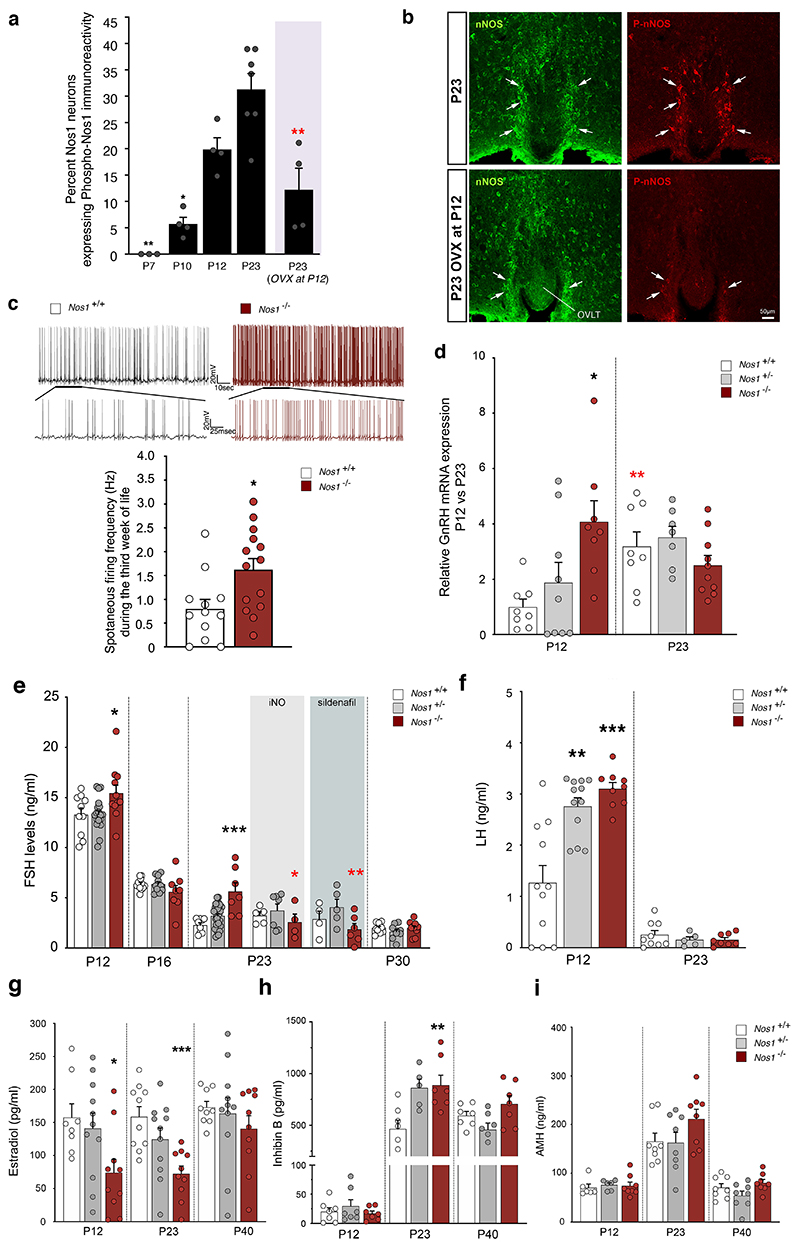
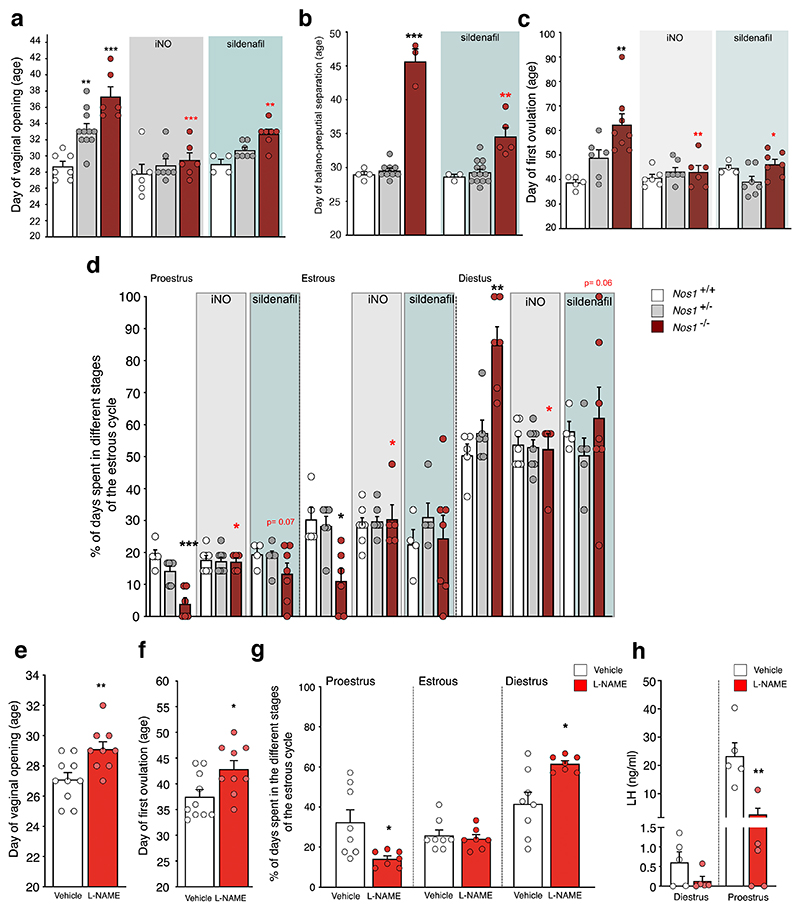
The nitric oxide (NO) signaling pathway in hypothalamic neurons plays a key role in the regulation of the secretion of gonadotropin-releasing hormone (GnRH), which is crucial for reproduction. We hypothesized that a disruption of neuronal NO synthase (NOS1) activity underlies some forms of hypogonadotropic hypogonadism. Whole-exome sequencing was performed on a cohort of 341 probands with congenital hypogonadotropic hypogonadism to identify ultrarare variants in NOS1. The activity of the identified NOS1 mutant proteins was assessed by their ability to promote nitrite and cGMP production in vitro. In addition, physiological and pharmacological characterization was carried out in a Nos1-deficient mouse model. We identified five heterozygous NOS1 loss-of-function mutations in six probands with congenital hypogonadotropic hypogonadism (2%), who displayed additional phenotypes including anosmia, hearing loss, and intellectual disability. NOS1 was found to be transiently expressed by GnRH neurons in the nose of both humans and mice, and Nos1 deficiency in mice resulted in dose-dependent defects in sexual maturation as well as in olfaction, hearing, and cognition. The pharmacological inhibition of NO production in postnatal mice revealed a critical time window during which Nos1 activity shaped minipuberty and sexual maturation. Inhaled NO treatment at minipuberty rescued both reproductive and behavioral phenotypes in Nos1-deficient mice. In summary, lack of NOS1 activity led to GnRH deficiency associated with sensory and intellectual comorbidities in humans and mice. NO treatment during minipuberty reversed deficits in sexual maturation, olfaction, and cognition in Nos1 mutant mice, suggesting a potential therapy for humans with NO deficiency.
Conflict of interest statement
Figures
References
-
- Boehm U, Bouloux PM, Dattani MT, de Roux N, Dode C, Dunkel L, Dwyer AA, Giacobini P, Hardelin JP, Juul A, Maghnie M, et al. Expert consensus document: European Consensus Statement on congenital hypogonadotropic hypogonadism--pathogenesis, diagnosis and treatment. Nat Rev Endocrinol. 2015;11:547–564. - PubMed
-
- Aydogan U, Aydogdu A, Akbulut H, Sonmez A, Yuksel S, Basaran Y, Uzun O, Bolu E, Saglam K. Increased frequency of anxiety, depression, quality of life and sexual life in young hypogonadotropic hypogonadal males and impacts of testosterone replacement therapy on these conditions. Endocr J. 2012;59:1099–1105. - PubMed
-
- Lasaite L, Ceponis J, Preiksa RT, Zilaitiene B. Impaired emotional state, quality of life and cognitive functions in young hypogonadal men. Andrologia. 2014;46:1107–1112. - PubMed
-
- Kuiri-Hanninen T, Sankilampi U, Dunkel L. Activation of the hypothalamic-pituitary-gonadal axis in infancy: minipuberty. Horm Res Paediatr. 2014;82:73–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
