

Opposing roles of hepatic stellate cell subpopulations in hepatocarcinogenesis
- PMID: 36198802
- PMCID: PMC9949942
- DOI: 10.1038/s41586-022-05289-6
Opposing roles of hepatic stellate cell subpopulations in hepatocarcinogenesis
Abstract
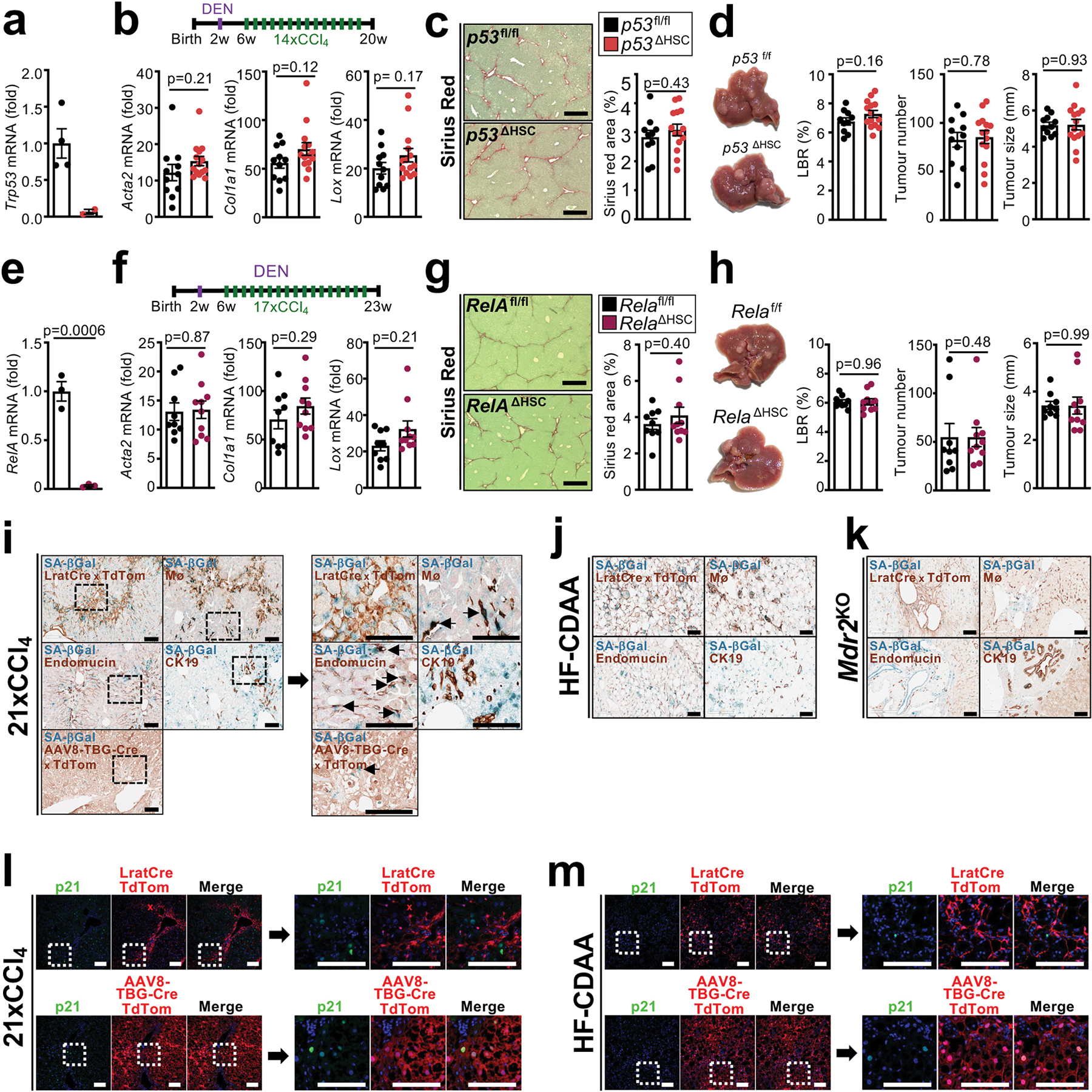
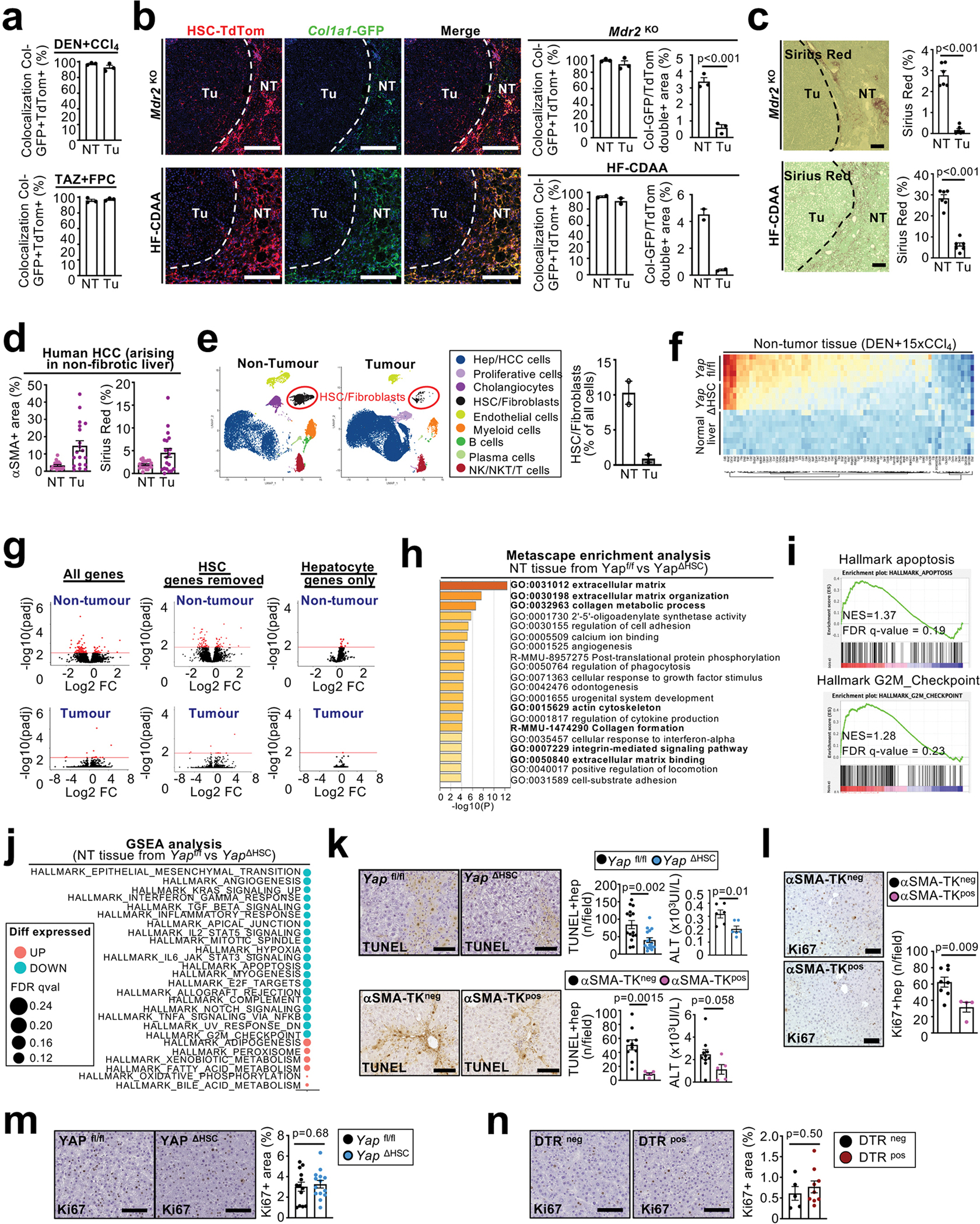
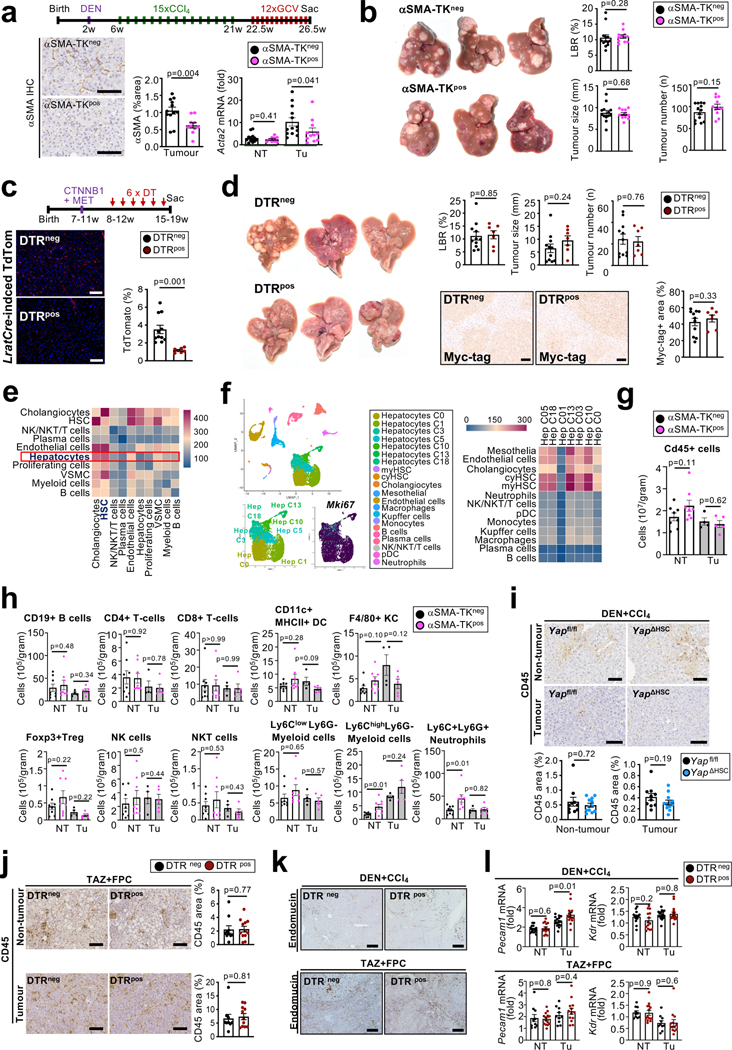
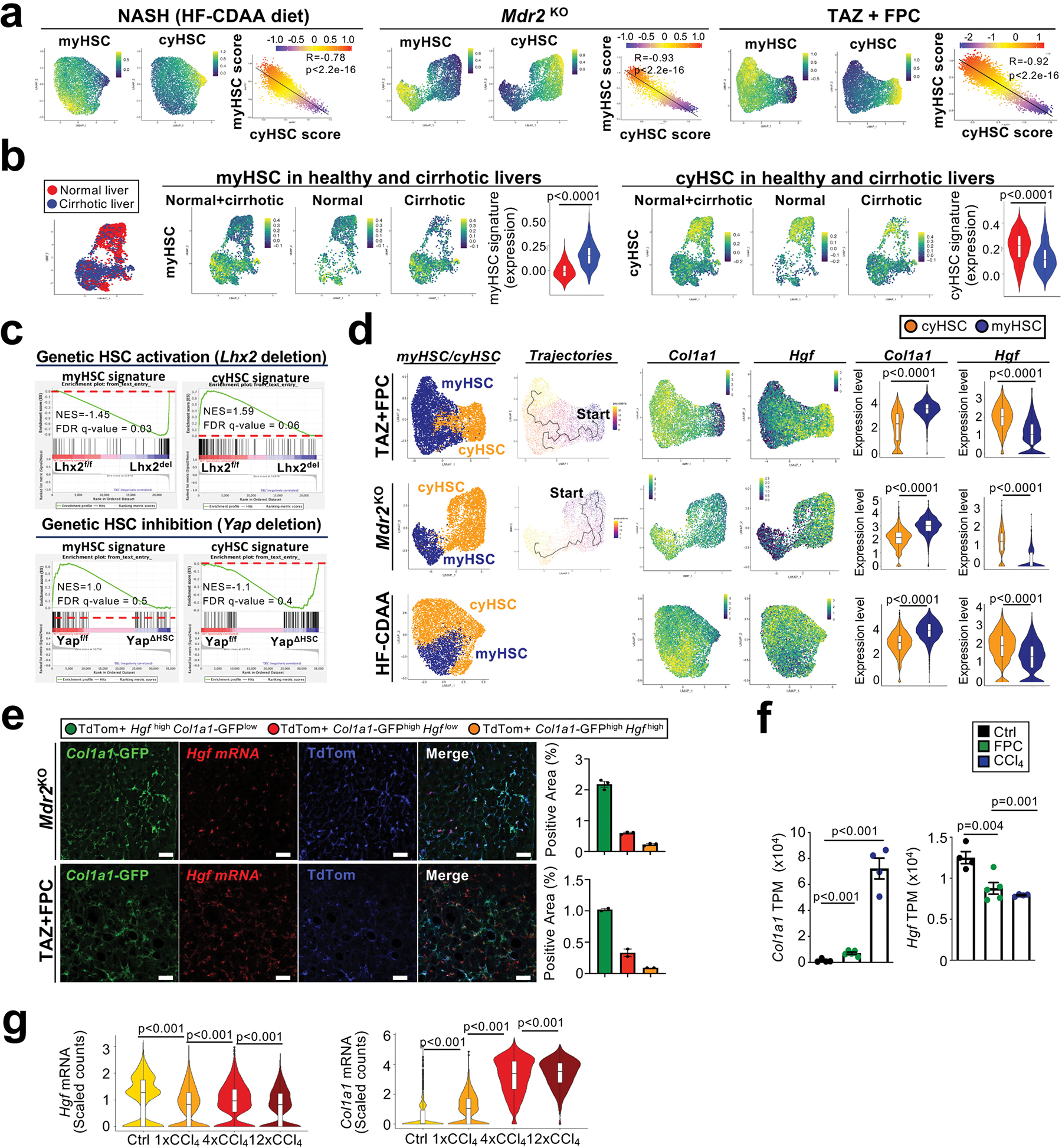
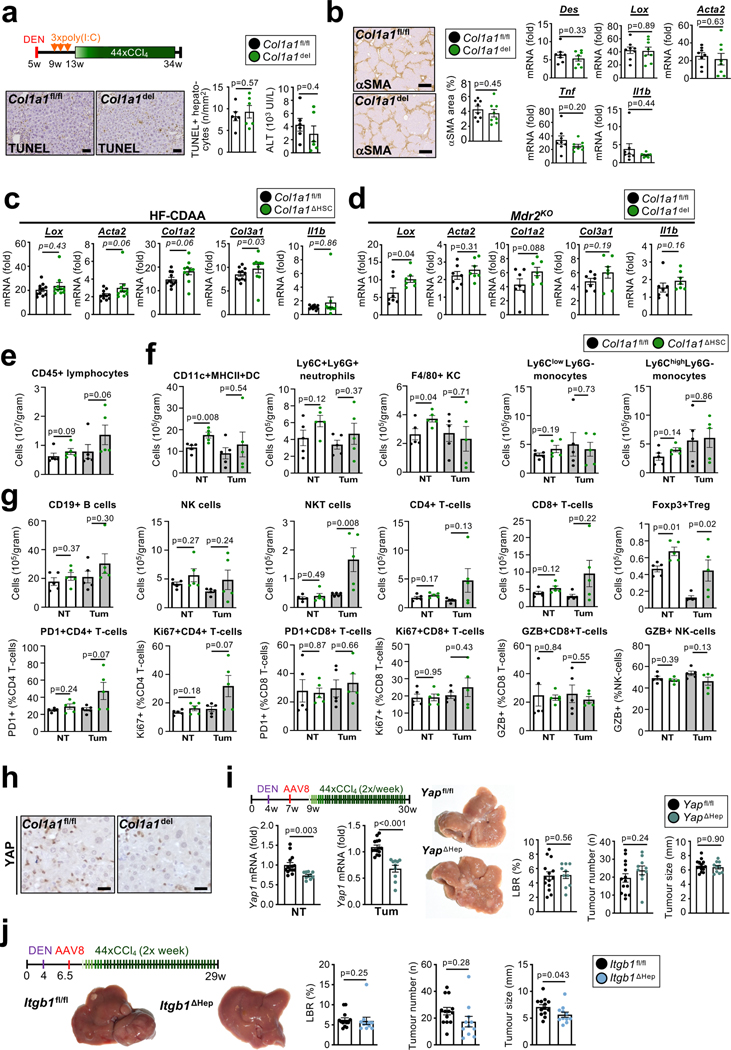
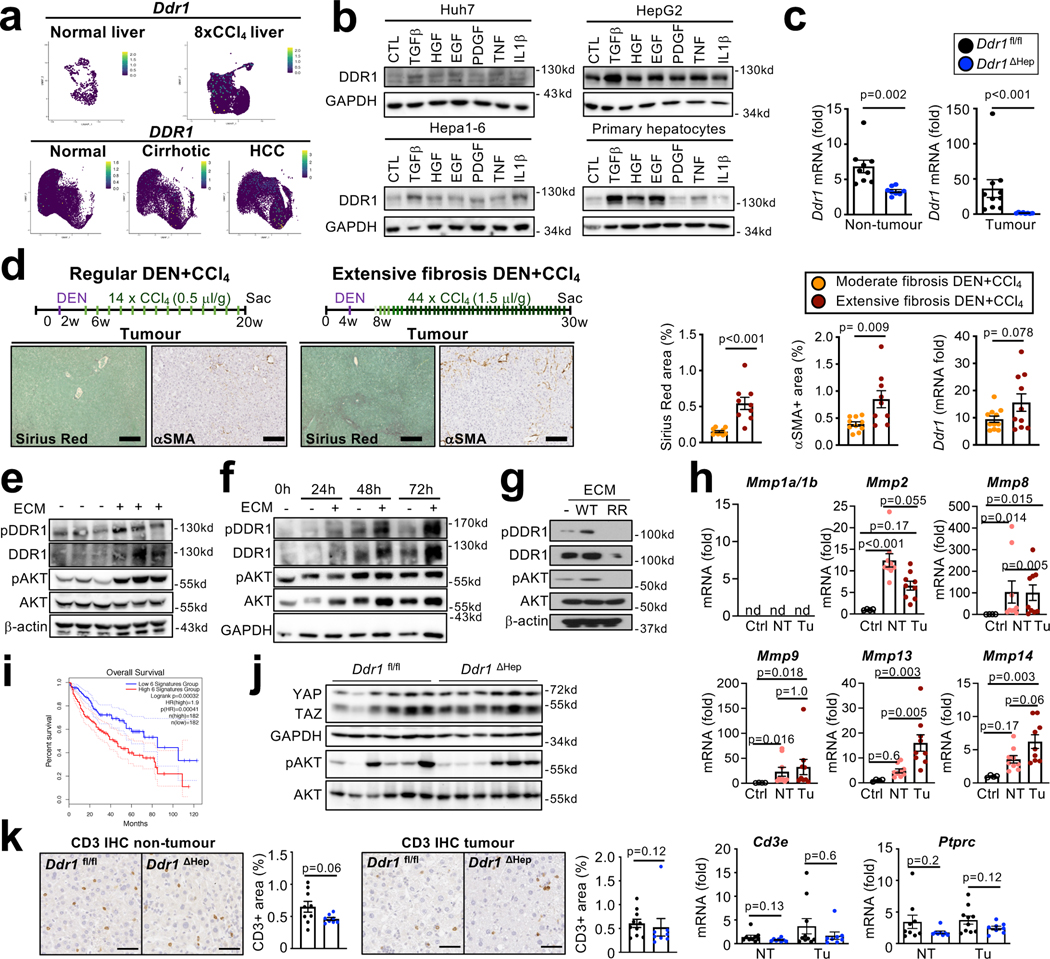
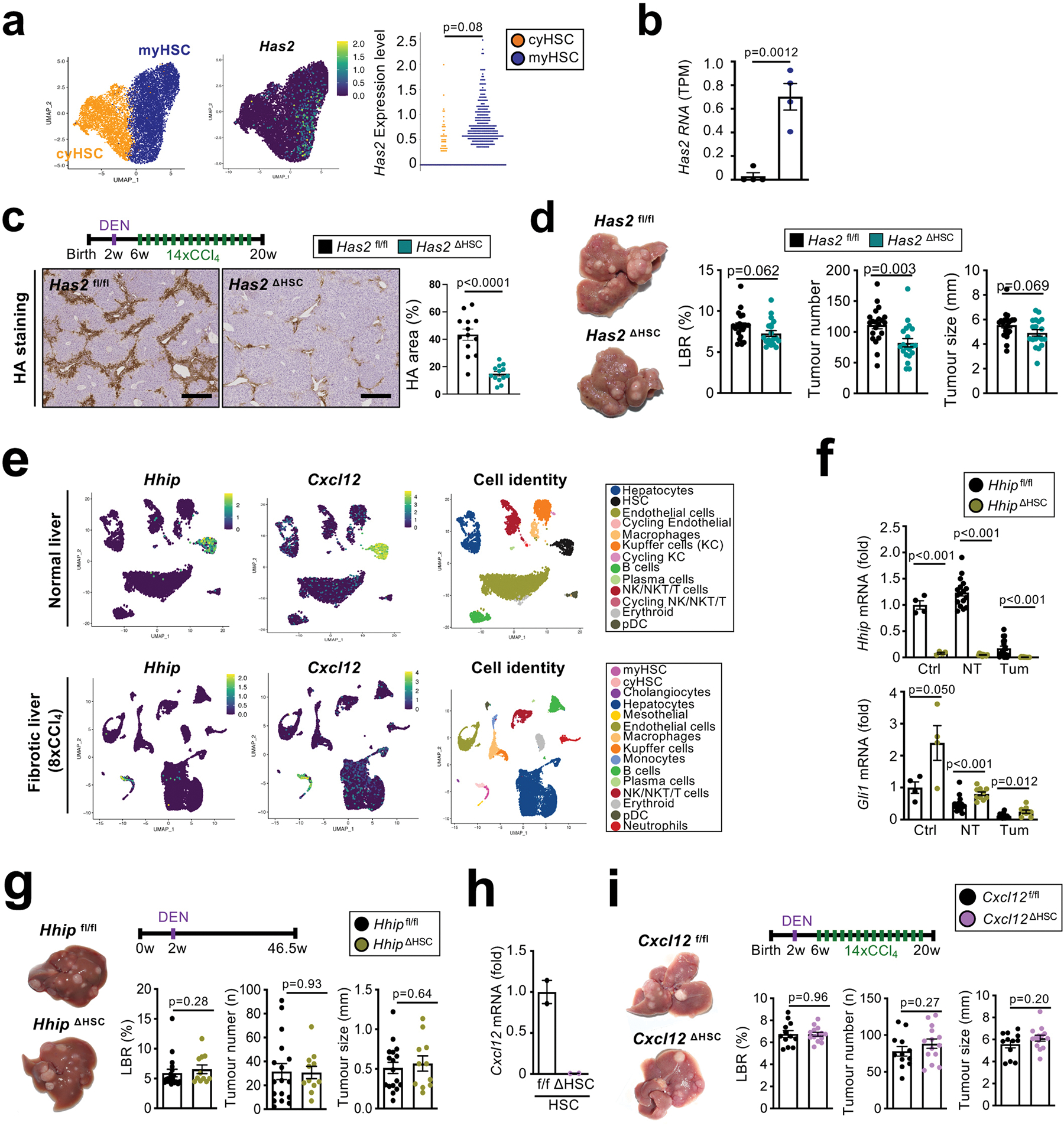
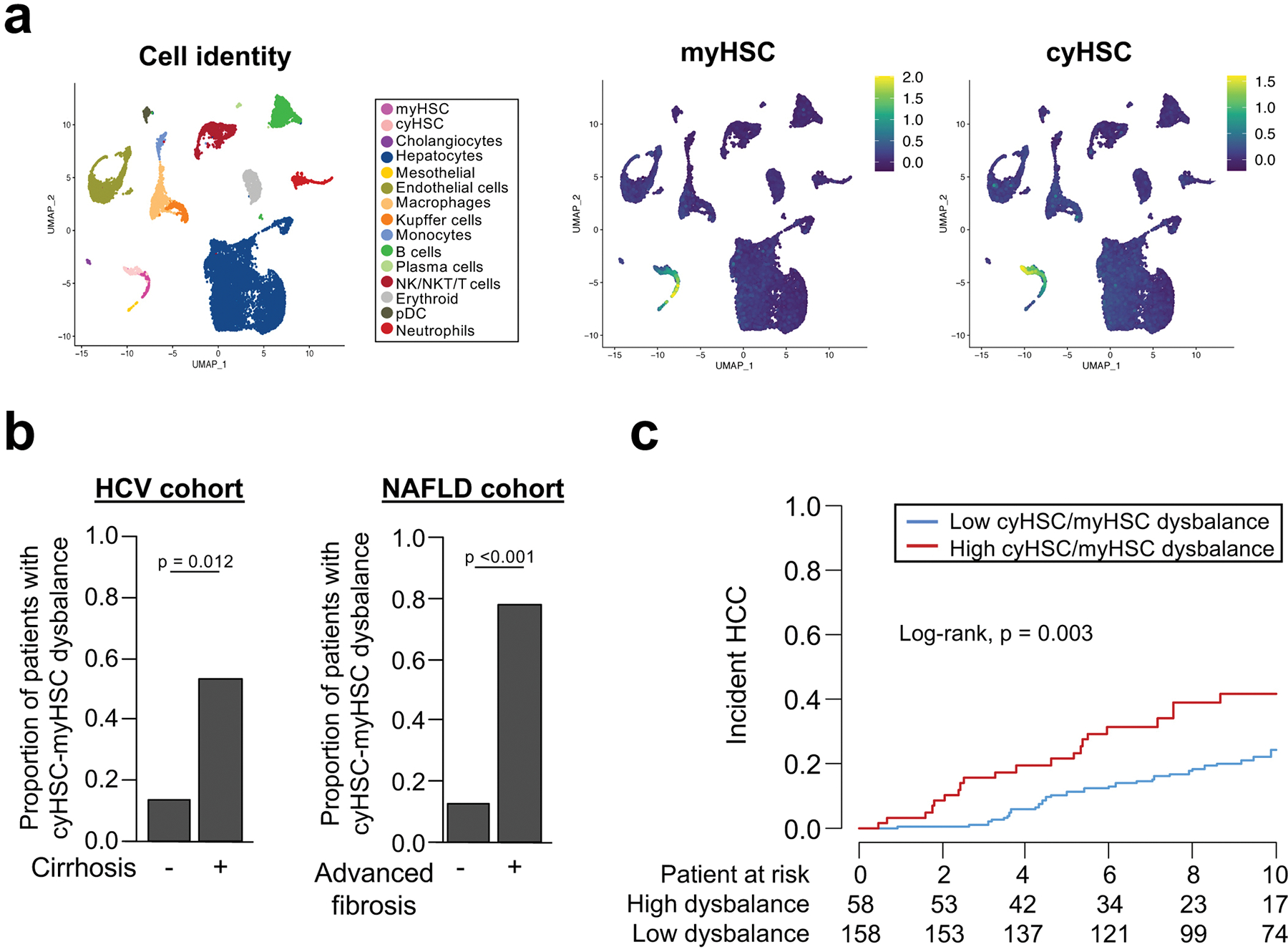
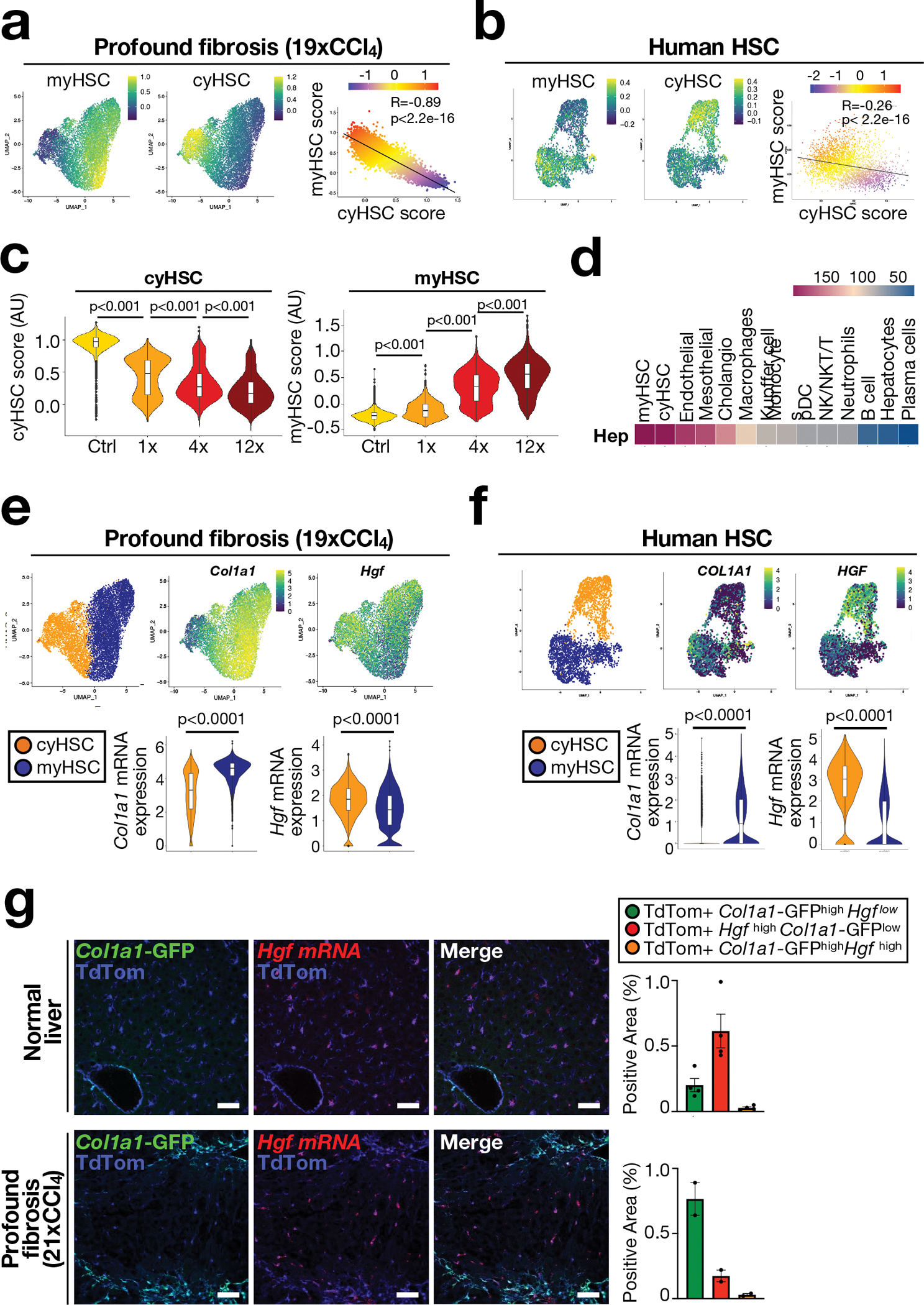
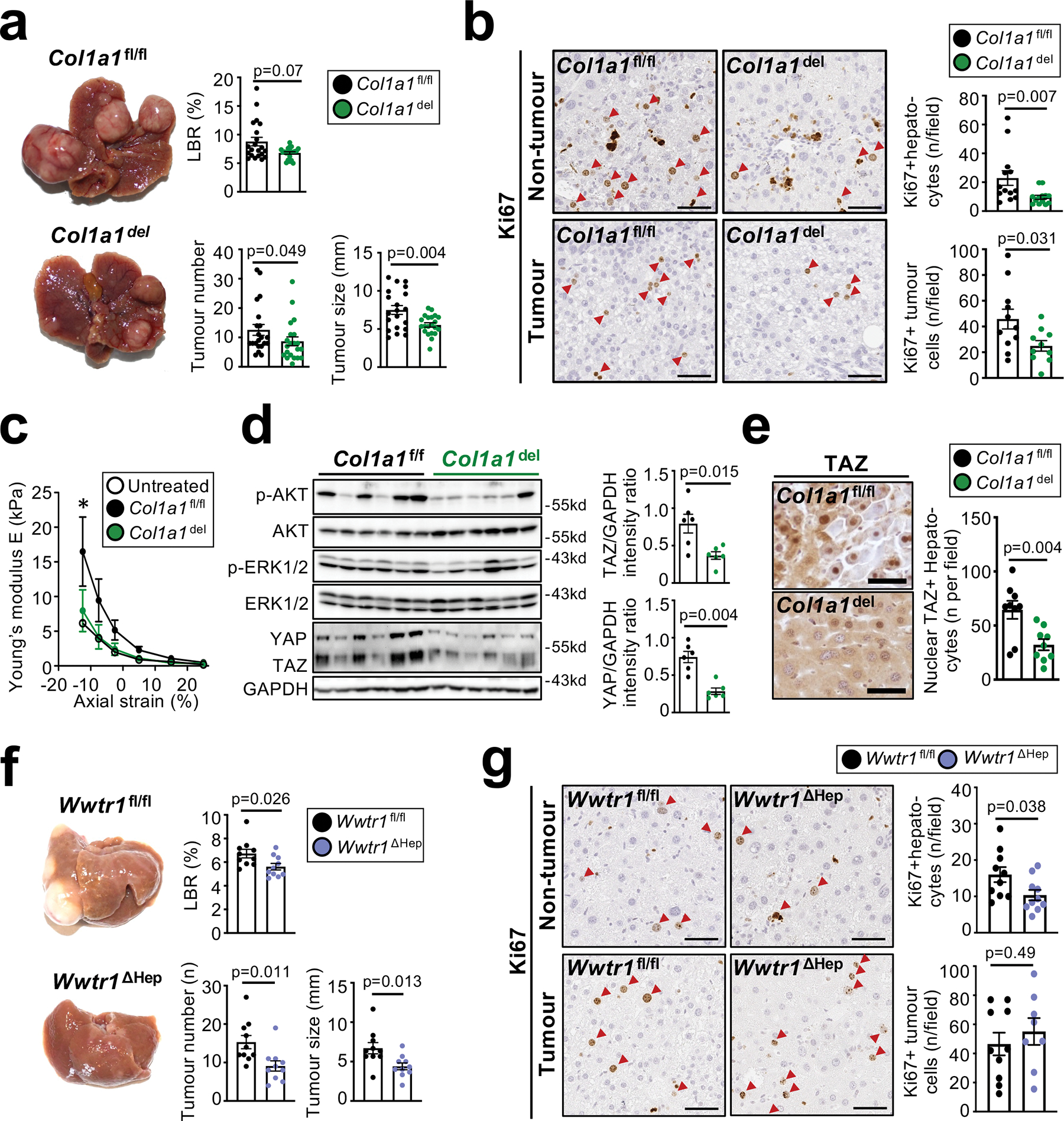
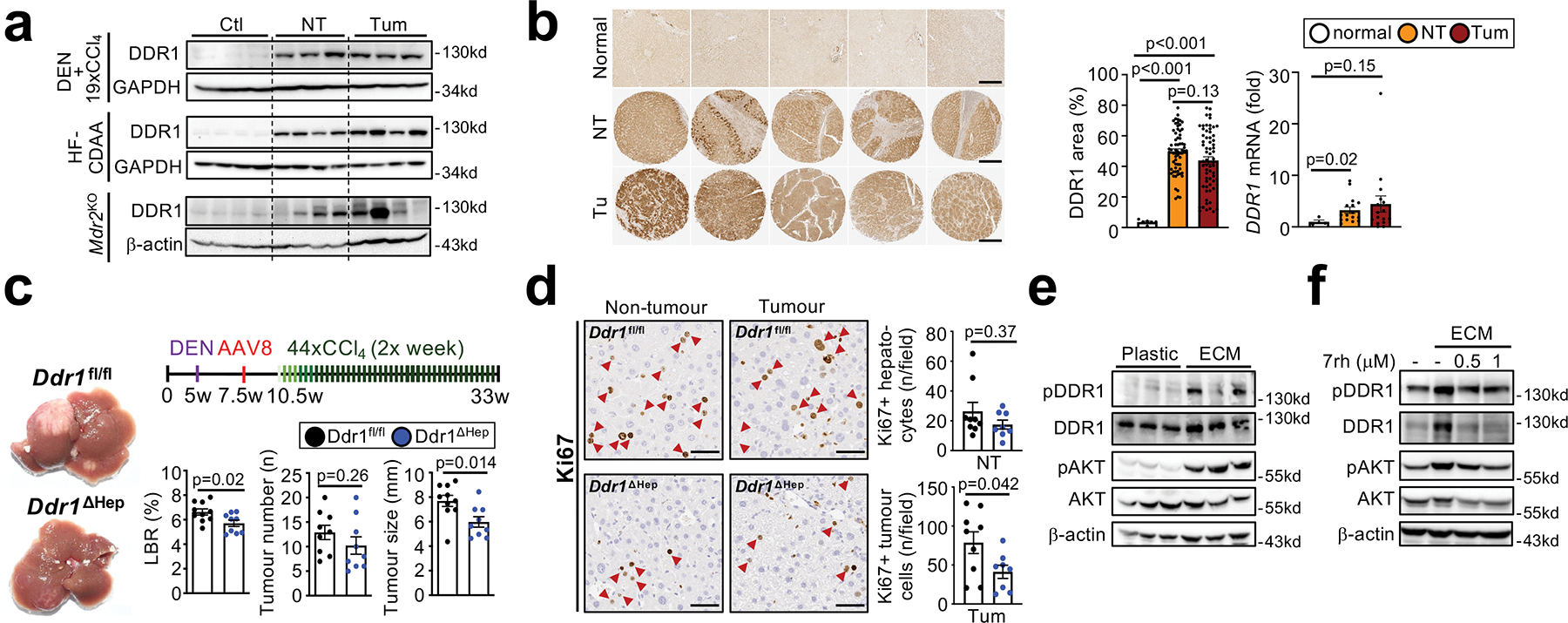
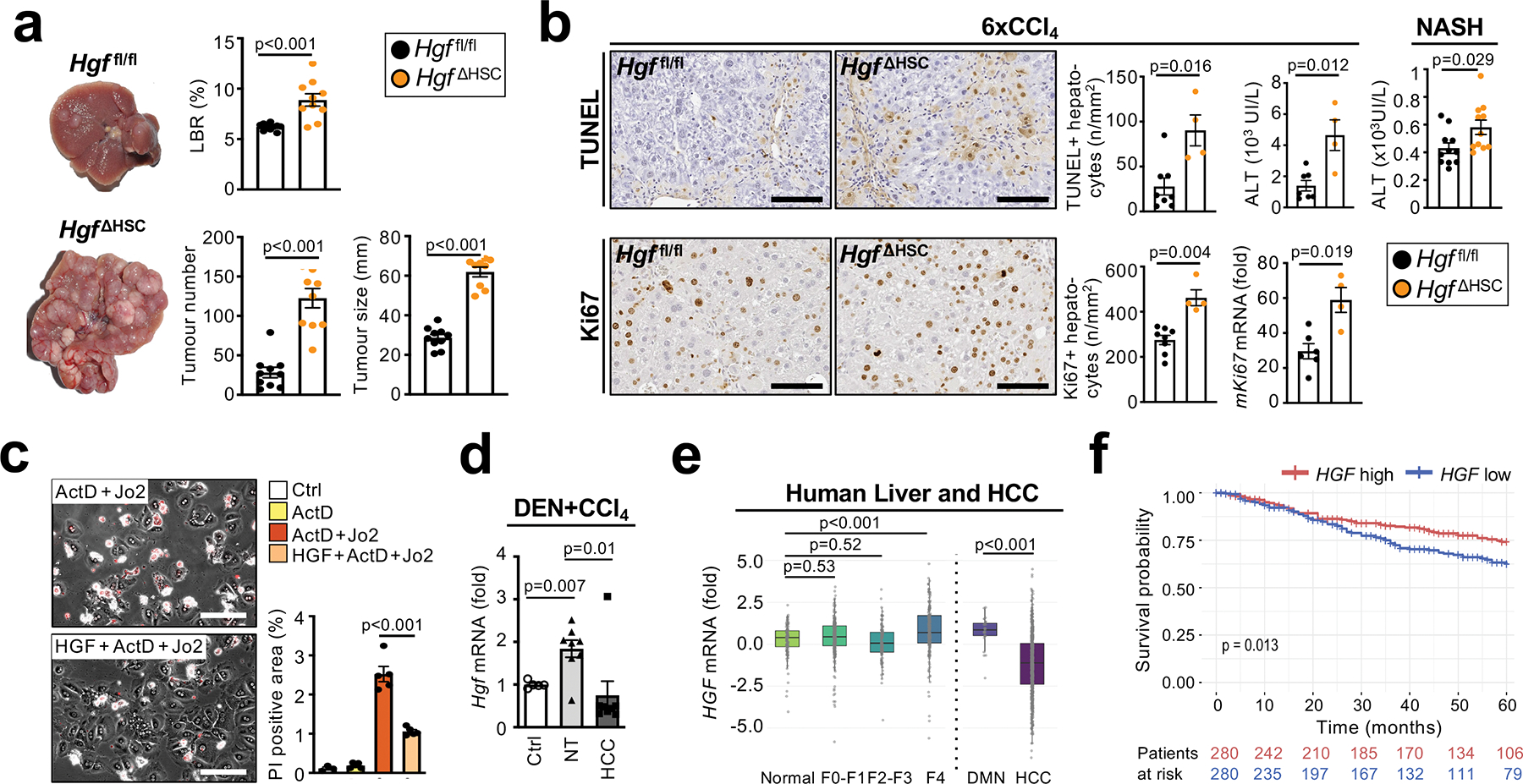
Hepatocellular carcinoma (HCC), the fourth leading cause of cancer mortality worldwide, develops almost exclusively in patients with chronic liver disease and advanced fibrosis1,2. Here we interrogated functions of hepatic stellate cells (HSCs), the main source of liver fibroblasts3, during hepatocarcinogenesis. Genetic depletion, activation or inhibition of HSCs in mouse models of HCC revealed their overall tumour-promoting role. HSCs were enriched in the preneoplastic environment, where they closely interacted with hepatocytes and modulated hepatocarcinogenesis by regulating hepatocyte proliferation and death. Analyses of mouse and human HSC subpopulations by single-cell RNA sequencing together with genetic ablation of subpopulation-enriched mediators revealed dual functions of HSCs in hepatocarcinogenesis. Hepatocyte growth factor, enriched in quiescent and cytokine-producing HSCs, protected against hepatocyte death and HCC development. By contrast, type I collagen, enriched in activated myofibroblastic HSCs, promoted proliferation and tumour development through increased stiffness and TAZ activation in pretumoural hepatocytes and through activation of discoidin domain receptor 1 in established tumours. An increased HSC imbalance between cytokine-producing HSCs and myofibroblastic HSCs during liver disease progression was associated with increased HCC risk in patients. In summary, the dynamic shift in HSC subpopulations and their mediators during chronic liver disease is associated with a switch from HCC protection to HCC promotion.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
COMPETING INTEREST DECLARATION
The authors declare no conflict of interest beside:
B.I. has received honoraria from consulting with Merck, J&J/Janssen Pharmaceuticals, AstraZeneca and Volastra Therapeutics.
M.K. is a founder and SAB member of Elgia Pharma and received research support from Merck, Janssen Gossamer4 Bio.
Figures







References
References (Methods)
MeSH terms
Substances
Grants and funding
- R21 CA263381/CA/NCI NIH HHS/United States
- F31 DK091980/DK/NIDDK NIH HHS/United States
- MRC_/Medical Research Council/United Kingdom
- 219542/Z/19/Z/WT_/Wellcome Trust/United Kingdom
- P30 CA013696/CA/NCI NIH HHS/United States
- R01 DK116620/DK/NIDDK NIH HHS/United States
- WT_/Wellcome Trust/United Kingdom
- R37 CA258829/CA/NCI NIH HHS/United States
- R01 DK128955/DK/NIDDK NIH HHS/United States
- P30 DK132710/DK/NIDDK NIH HHS/United States
- R01 CA211794/CA/NCI NIH HHS/United States
- R01 CA190844/CA/NCI NIH HHS/United States
- S10 OD020056/OD/NIH HHS/United States
- R01 CA228483/CA/NCI NIH HHS/United States
- R01 CA233794/CA/NCI NIH HHS/United States
- R01 CA234128/CA/NCI NIH HHS/United States
- U01 CA274295/CA/NCI NIH HHS/United States
- R01 CA262424/CA/NCI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
