

Nitrate-responsive OsMADS27 promotes salt tolerance in rice
- PMID: 36199247
- PMCID: PMC10030316
- DOI: 10.1016/j.xplc.2022.100458
Nitrate-responsive OsMADS27 promotes salt tolerance in rice
Abstract
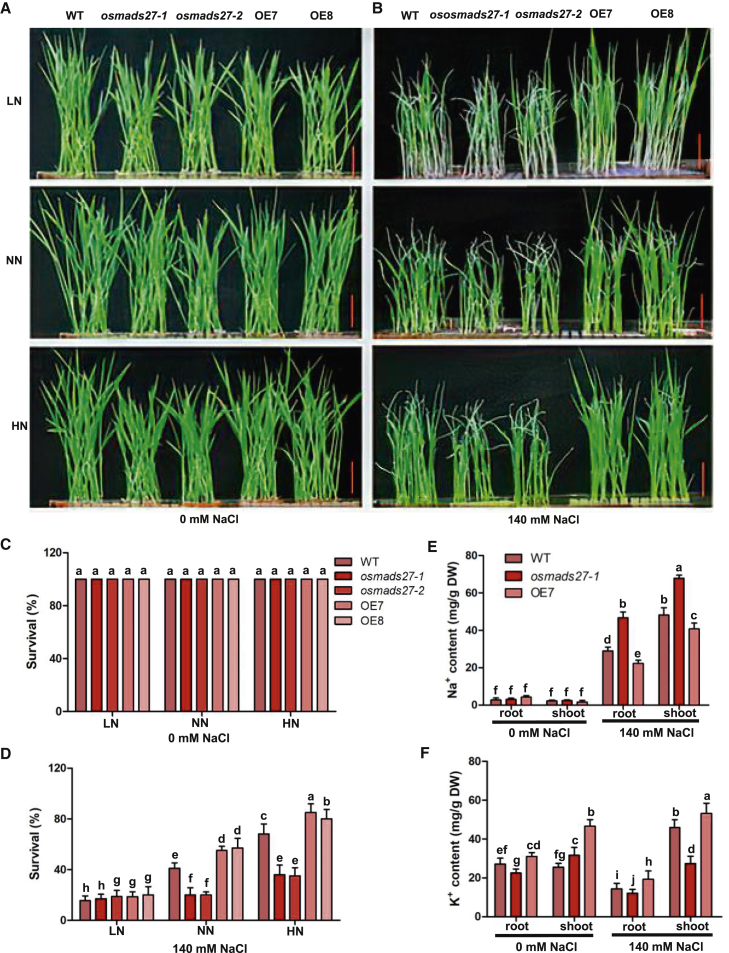
Salt stress is a major constraint on plant growth and yield. Nitrogen (N) fertilizers are known to alleviate salt stress. However, the underlying molecular mechanisms remain unclear. Here, we show that nitrate-dependent salt tolerance is mediated by OsMADS27 in rice. The expression of OsMADS27 is specifically induced by nitrate. The salt-inducible expression of OsMADS27 is also nitrate dependent. OsMADS27 knockout mutants are more sensitive to salt stress than the wild type, whereas OsMADS27 overexpression lines are more tolerant. Transcriptomic analyses revealed that OsMADS27 upregulates the expression of a number of known stress-responsive genes as well as those involved in ion homeostasis and antioxidation. We demonstrate that OsMADS27 directly binds to the promoters of OsHKT1.1 and OsSPL7 to regulate their expression. Notably, OsMADS27-mediated salt tolerance is nitrate dependent and positively correlated with nitrate concentration. Our results reveal the role of nitrate-responsive OsMADS27 and its downstream target genes in salt tolerance, providing a molecular mechanism for the enhancement of salt tolerance by nitrogen fertilizers in rice. OsMADS27 overexpression increased grain yield under salt stress in the presence of sufficient nitrate, suggesting that OsMADS27 is a promising candidate for the improvement of salt tolerance in rice.
Keywords: OsMADS27; grain yield; nitrate-dependent salt tolerance; salt stress.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Figures






Similar articles
-
Nuclear translocation of OsMADS25 facilitated by OsNAR2.1 in reponse to nitrate signals promotes rice root growth by targeting OsMADS27 and OsARF7.Plant Commun. 2023 Nov 13;4(6):100642. doi: 10.1016/j.xplc.2023.100642. Epub 2023 Jun 24. Plant Commun. 2023. PMID: 37353931 Free PMC article.
-
OsMADS27 regulates the root development in a NO3--Dependent manner and modulates the salt tolerance in rice (Oryza sativa L.).Plant Sci. 2018 Dec;277:20-32. doi: 10.1016/j.plantsci.2018.09.004. Epub 2018 Sep 10. Plant Sci. 2018. PMID: 30466586
-
Nitrate-dependent regulation of miR444-OsMADS27 signalling cascade controls root development in rice.J Exp Bot. 2022 Jun 2;73(11):3511-3530. doi: 10.1093/jxb/erac083. J Exp Bot. 2022. PMID: 35243491
-
Advances in Sensing, Response and Regulation Mechanism of Salt Tolerance in Rice.Int J Mol Sci. 2021 Feb 24;22(5):2254. doi: 10.3390/ijms22052254. Int J Mol Sci. 2021. PMID: 33668247 Free PMC article. Review.
-
Root Development and Stress Tolerance in rice: The Key to Improving Stress Tolerance without Yield Penalties.Int J Mol Sci. 2020 Mar 6;21(5):1807. doi: 10.3390/ijms21051807. Int J Mol Sci. 2020. PMID: 32155710 Free PMC article. Review.
Cited by
-
Genome-wide identification, characterization, and expression analysis of the MADS-box gene family in grass pea (Lathyrus sativus) under salt stress conditions.BMC Genomics. 2025 May 21;26(1):519. doi: 10.1186/s12864-025-11661-3. BMC Genomics. 2025. PMID: 40399794 Free PMC article.
-
Finding Balance in Adversity: Nitrate Signaling as the Key to Plant Growth, Resilience, and Stress Response.Int J Mol Sci. 2023 Sep 22;24(19):14406. doi: 10.3390/ijms241914406. Int J Mol Sci. 2023. PMID: 37833854 Free PMC article. Review.
-
Sustaining nitrogen dynamics: A critical aspect for improving salt tolerance in plants.Front Plant Sci. 2023 Feb 23;14:1087946. doi: 10.3389/fpls.2023.1087946. eCollection 2023. Front Plant Sci. 2023. PMID: 36909406 Free PMC article. Review.
-
Genome-wide analysis of MADS-box transcription factor gene family in wild emmer wheat (Triticum turgidum subsp. dicoccoides).PLoS One. 2024 Mar 7;19(3):e0300159. doi: 10.1371/journal.pone.0300159. eCollection 2024. PLoS One. 2024. PMID: 38451993 Free PMC article.
-
Nuclear translocation of OsMADS25 facilitated by OsNAR2.1 in reponse to nitrate signals promotes rice root growth by targeting OsMADS27 and OsARF7.Plant Commun. 2023 Nov 13;4(6):100642. doi: 10.1016/j.xplc.2023.100642. Epub 2023 Jun 24. Plant Commun. 2023. PMID: 37353931 Free PMC article.
References
-
- Ahammed G.J., Li X., Yang Y., Liu C., Zhou G., Wan H., Cheng Y. Tomato WRKY81 acts as a negative regulator for drought tolerance by modulating guard cell H2O2–mediated stomatal closure. Environ. Exp. Bot. 2020;171:103960.
-
- Aragão R.M., Silva E.N., Vieira C.F., Silveira J.A.G. High supply of NO 3− mitigates salinity effects through an enhancement in the efficiency of photosystem II and CO 2 assimilation in Jatropha curcas plants. Acta Physiol. Plant. 2012;34:2135–2143.
-
- Asano T., Hayashi N., Kobayashi M., Aoki N., Miyao A., Mitsuhara I., Ichikawa H., Komatsu S., Hirochika H., Kikuchi S., et al. A rice calcium-dependent protein kinase OsCPK12 oppositely modulates salt-stress tolerance and blast disease resistance. Plant J. 2012;69:26–36. - PubMed
-
- Ashraf M., Athar H., Harris P., Kwon T. Some prospective strategies for improving crop salt tolerance. Adv. Agron. 2008;97:45–110.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
