

RAD21 amplification epigenetically suppresses interferon signaling to promote immune evasion in ovarian cancer
- PMID: 36201246
- PMCID: PMC9663158
- DOI: 10.1172/JCI159628
RAD21 amplification epigenetically suppresses interferon signaling to promote immune evasion in ovarian cancer
Abstract
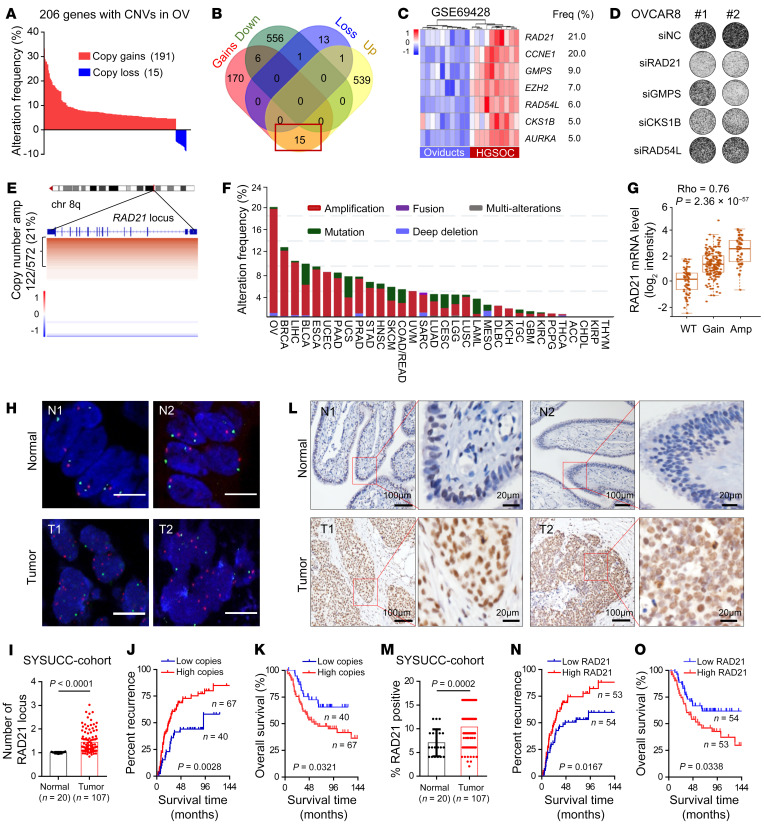
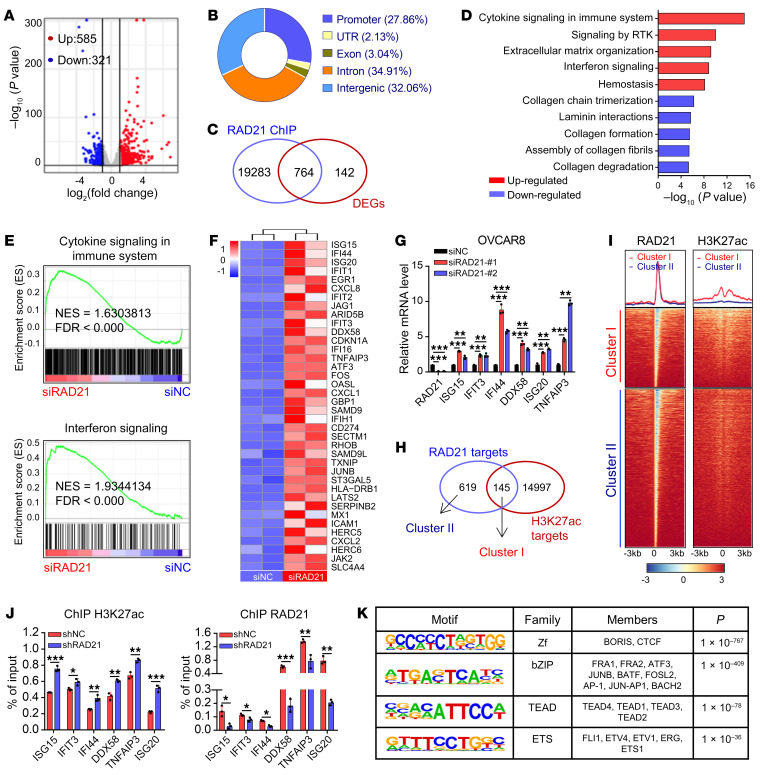
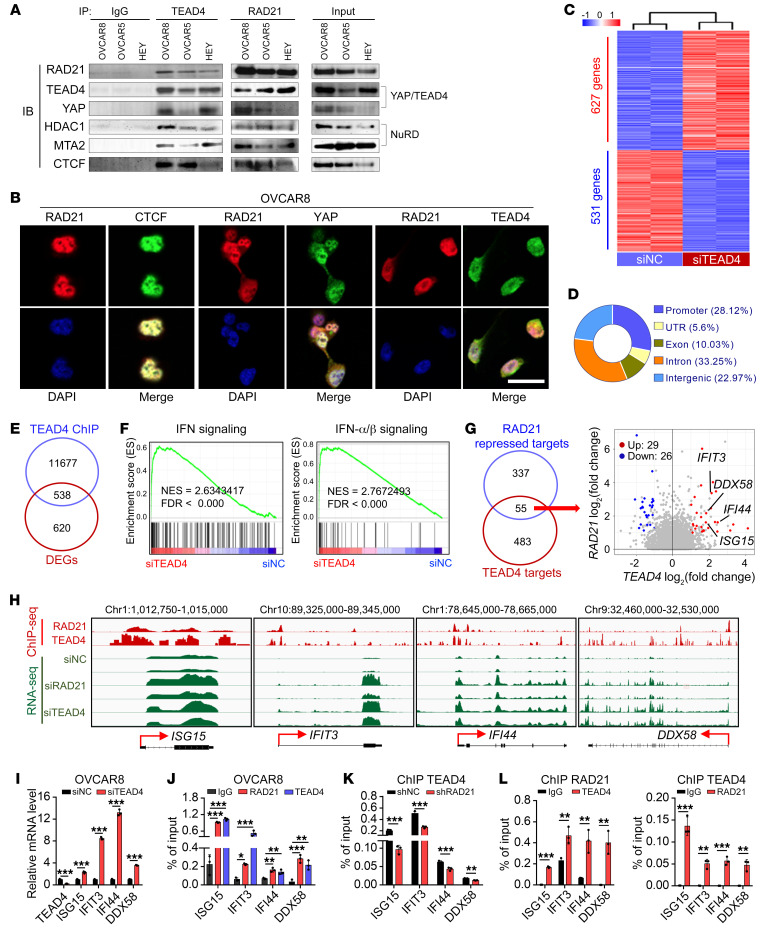
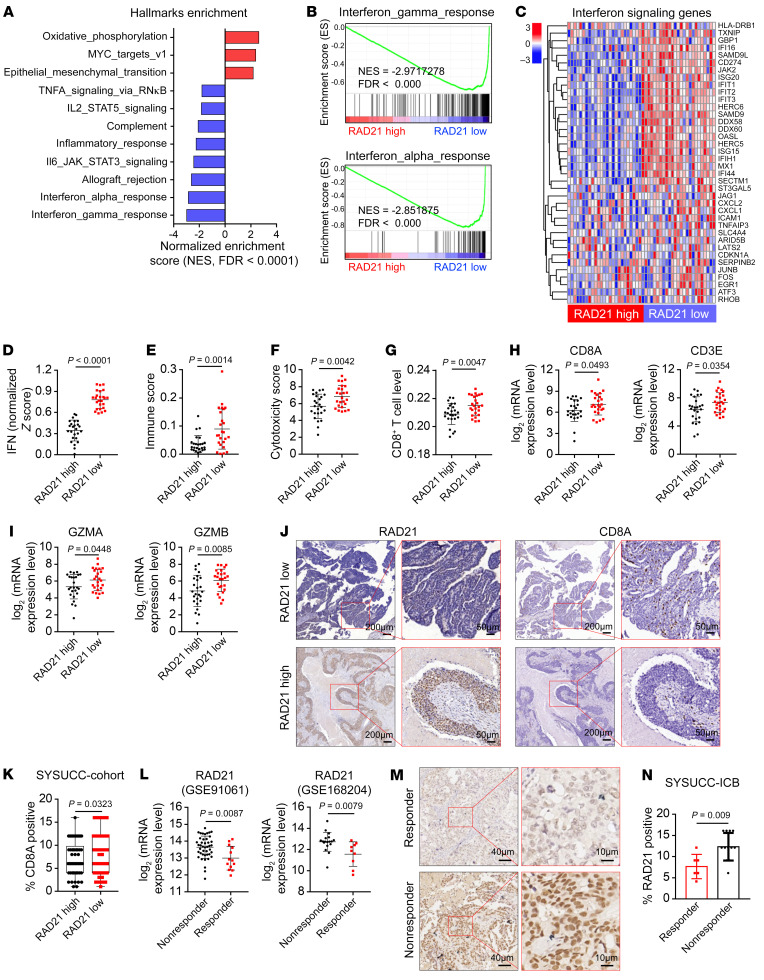
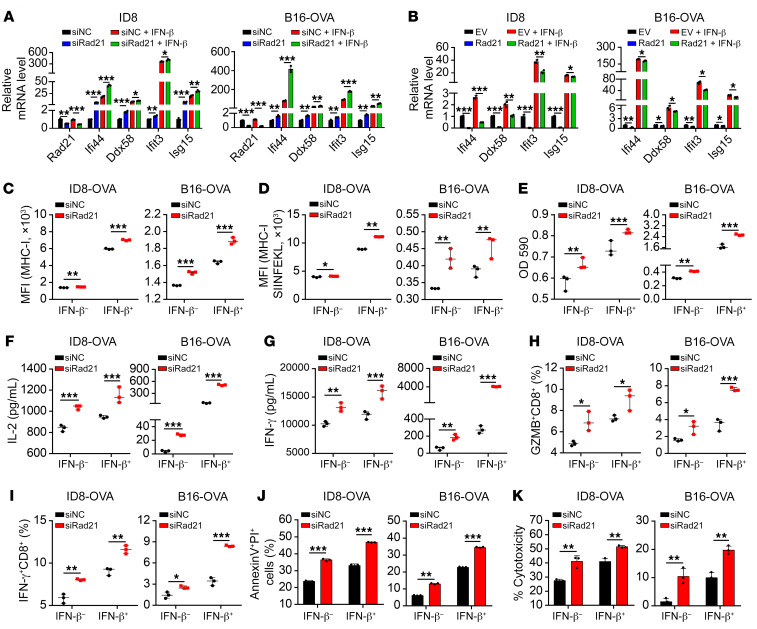
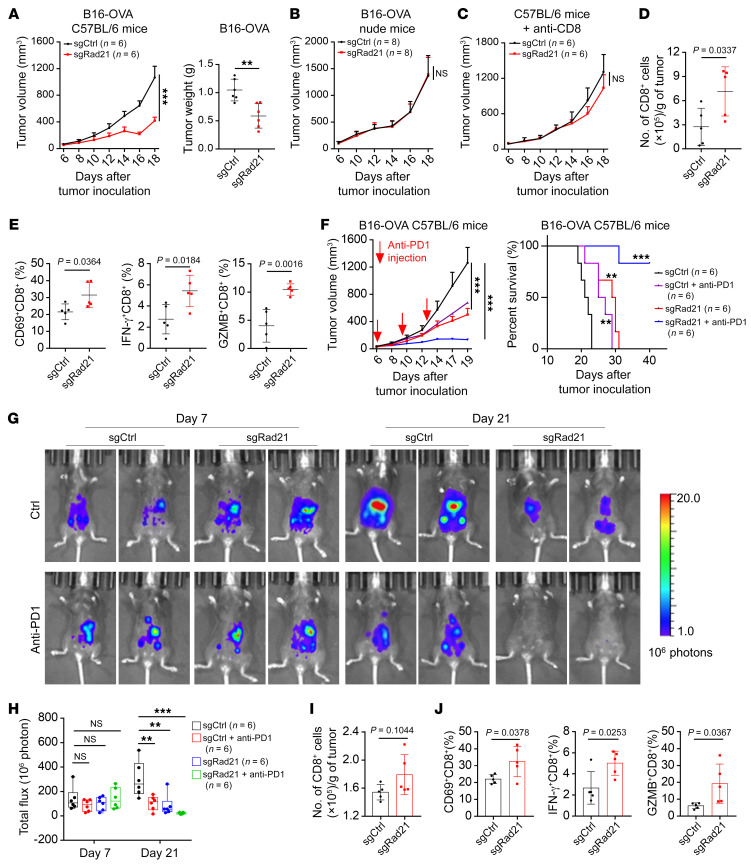
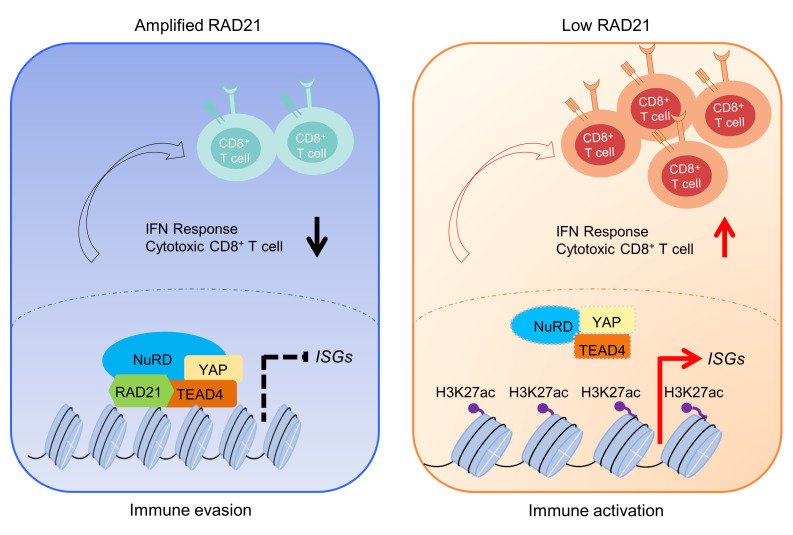
Prevalent copy number alteration is the most prominent genetic characteristic associated with ovarian cancer (OV) development, but its role in immune evasion has not been fully elucidated. In this study, we identified RAD21, a key component of the cohesin complex, as a frequently amplified oncogene that could modulate immune response in OV. Through interrogating the RAD21-regulated transcriptional program, we found that RAD21 directly interacts with YAP/TEAD4 transcriptional corepressors and recruits the NuRD complex to suppress interferon (IFN) signaling. In multiple clinical cohorts, RAD21 overexpression is inversely correlated with IFN signature gene expression in OV. We further demonstrated in murine syngeneic tumor models that RAD21 ablation potentiated anti-PD-1 efficacy with increased intratumoral CD8+ T cell effector activity. Our study identifies a RAD21-YAP/TEAD4-NuRD corepressor complex in immune modulation, and thus provides a potential target and biomarker for precision immunotherapy in OV.
Keywords: Cancer immunotherapy; Epigenetics; Genetic instability; Immunology; Oncology.
Conflict of interest statement
Figures
References
-
- Wei SC, et al. Fundamental mechanisms of immune checkpoint blockade therapy. Cancer Discov. 2018;8(9):1069–1086. doi: 10.1158/2159-8290.CD-18-0367. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
