

Lysosomal lipid alterations caused by glucocerebrosidase deficiency promote lysosomal dysfunction, chaperone-mediated-autophagy deficiency, and alpha-synuclein pathology
- PMID: 36202848
- PMCID: PMC9537323
- DOI: 10.1038/s41531-022-00397-6
Lysosomal lipid alterations caused by glucocerebrosidase deficiency promote lysosomal dysfunction, chaperone-mediated-autophagy deficiency, and alpha-synuclein pathology
Abstract
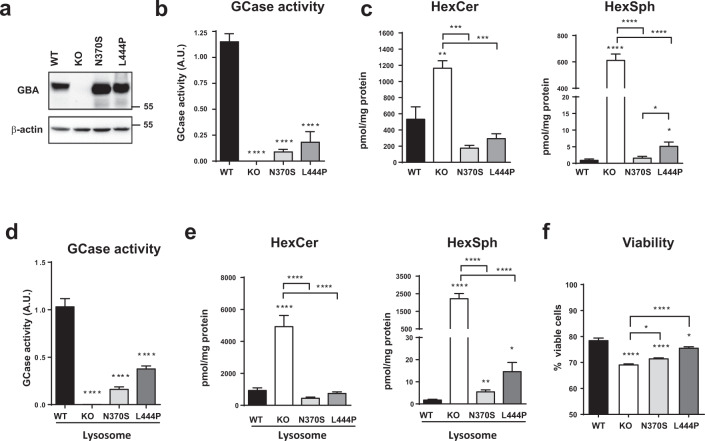
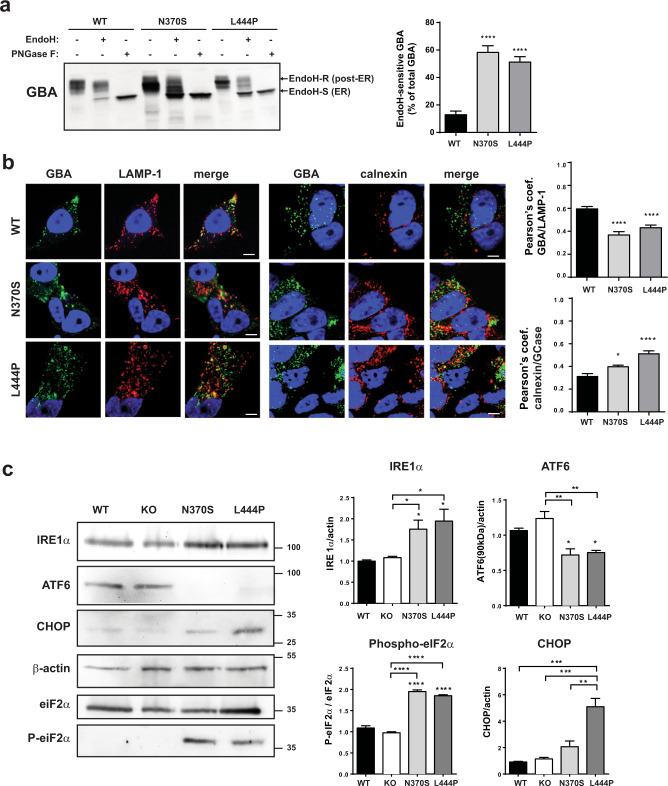
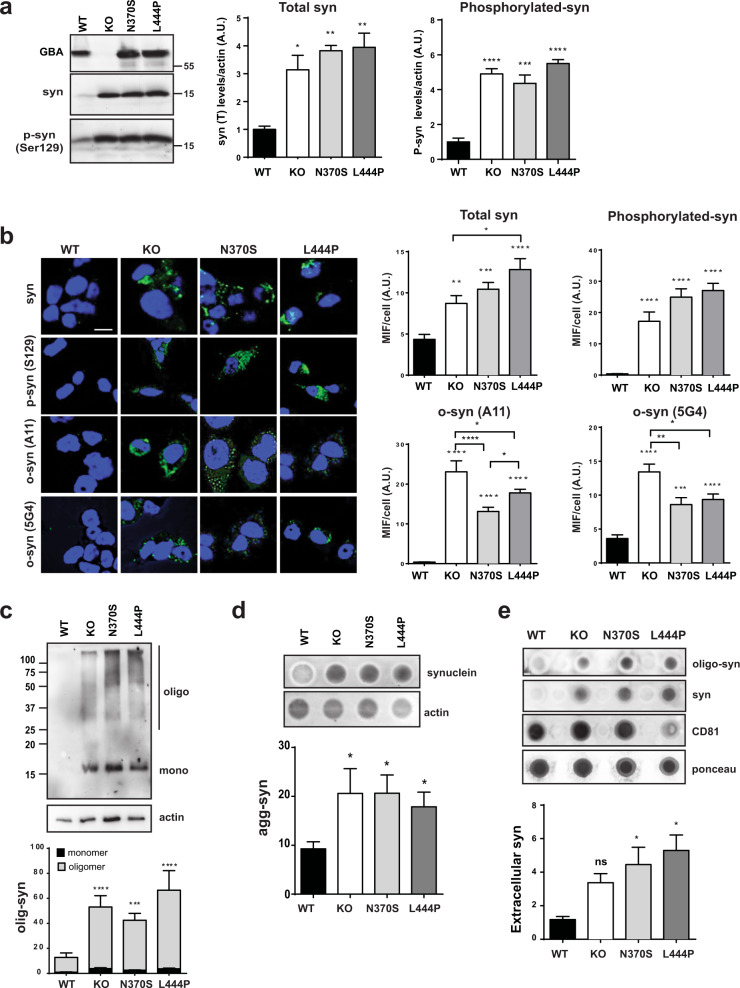
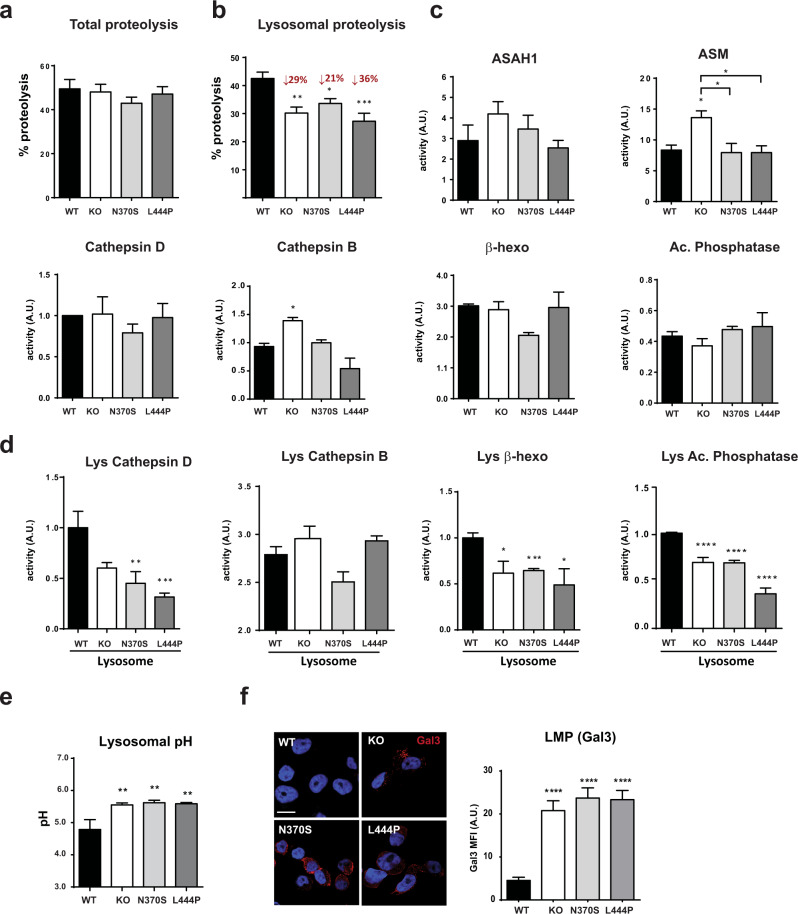
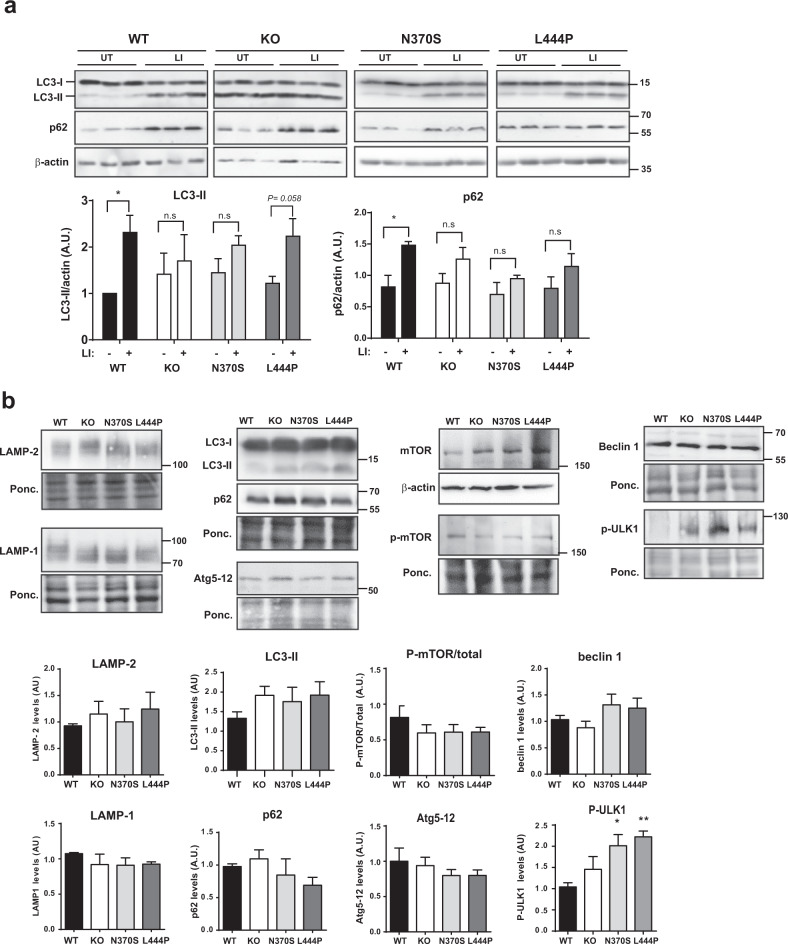
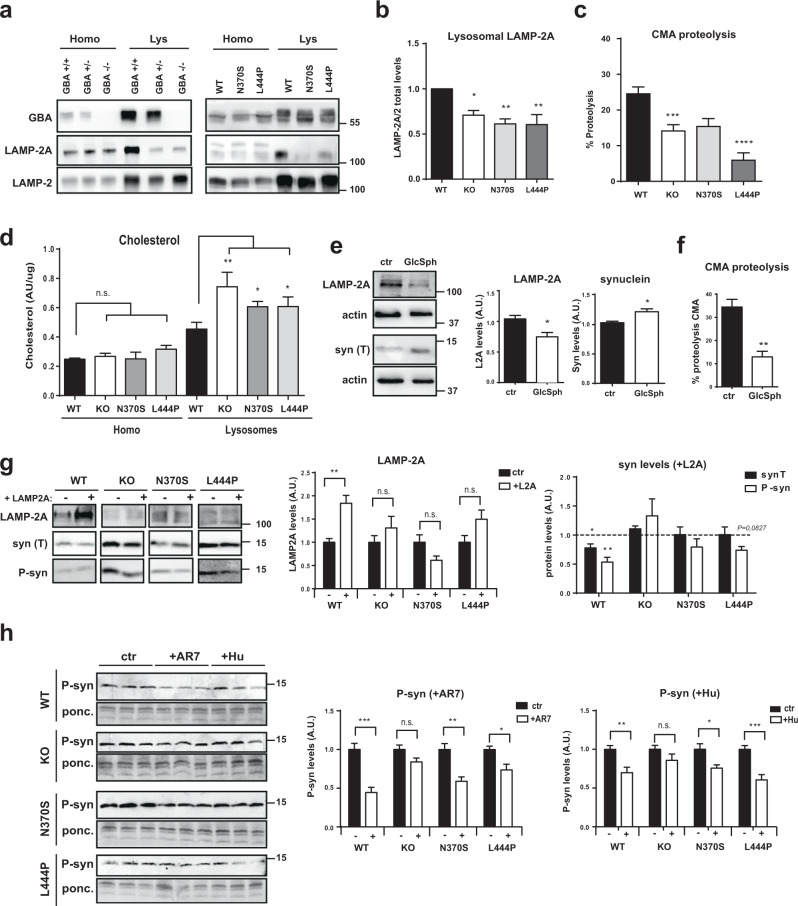
Mutations in the GBA gene that encodes the lysosomal enzyme β-glucocerebrosidase (GCase) are a major genetic risk factor for Parkinson's disease (PD). In this study, we generated a set of differentiated and stable human dopaminergic cell lines that express the two most prevalent GBA mutations as well as GBA knockout cell lines as a in vitro disease modeling system to study the relationship between mutant GBA and the abnormal accumulation of α-synuclein. We performed a deep analysis of the consequences triggered by the presence of mutant GBA protein and the loss of GCase activity in different cellular compartments, focusing primarily on the lysosomal compartment, and analyzed in detail the lysosomal activity, composition, and integrity. The loss of GCase activity generates extensive lysosomal dysfunction, promoting the loss of activity of other lysosomal enzymes, affecting lysosomal membrane stability, promoting intralysosomal pH changes, and favoring the intralysosomal accumulation of sphingolipids and cholesterol. These local events, occurring only at a subcellular level, lead to an impairment of autophagy pathways, particularly chaperone-mediated autophagy, the main α-synuclein degradative pathway. The findings of this study highlighted the role of lysosomal function and lipid metabolism in PD and allowed us to describe a molecular mechanism to understand how mutations in GBA can contribute to an abnormal accumulation of different α-synuclein neurotoxic species in PD pathology.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
Grants and funding
- PI20/00728/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- PI17/00496/Ministry of Economy and Competitiveness | Instituto de Salud Carlos III (Institute of Health Carlos III)
- MJFF 16182/Michael J. Fox Foundation for Parkinson's Research (Michael J. Fox Foundation)
- NANOERT/Fundación BBVA (BBVA Foundation)
- SLT006/17/00268/Government of Catalonia | Departament de Salut, Generalitat de Catalunya
LinkOut - more resources
Full Text Sources
