

Spatiotemporal dynamics of membrane surface charge regulates cell polarity and migration
- PMID: 36202973
- PMCID: PMC10029748
- DOI: 10.1038/s41556-022-00997-7
Spatiotemporal dynamics of membrane surface charge regulates cell polarity and migration
Abstract
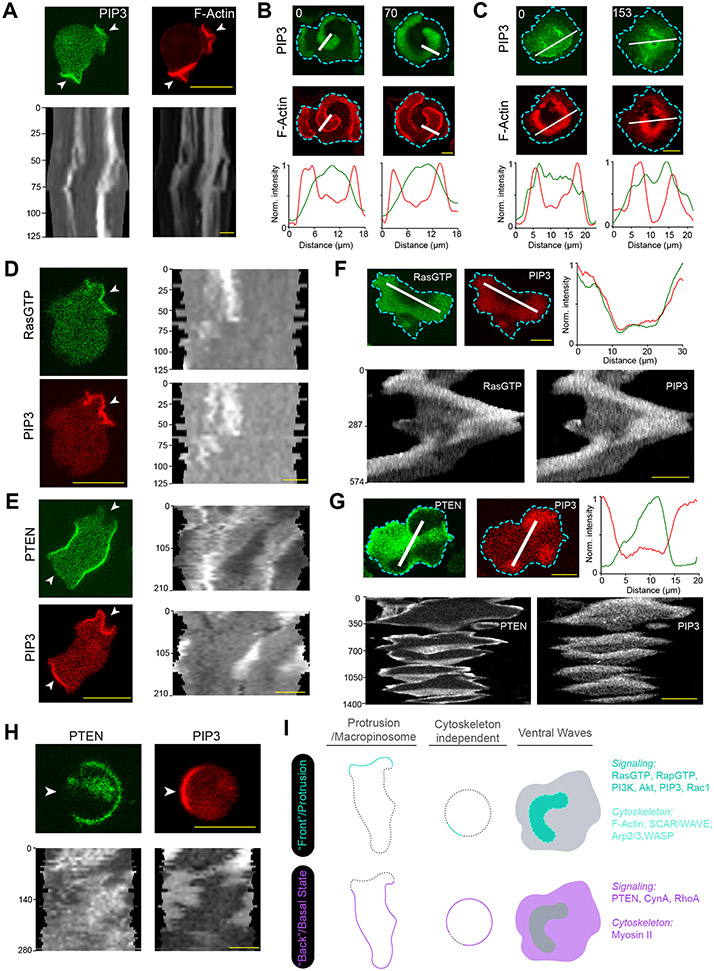
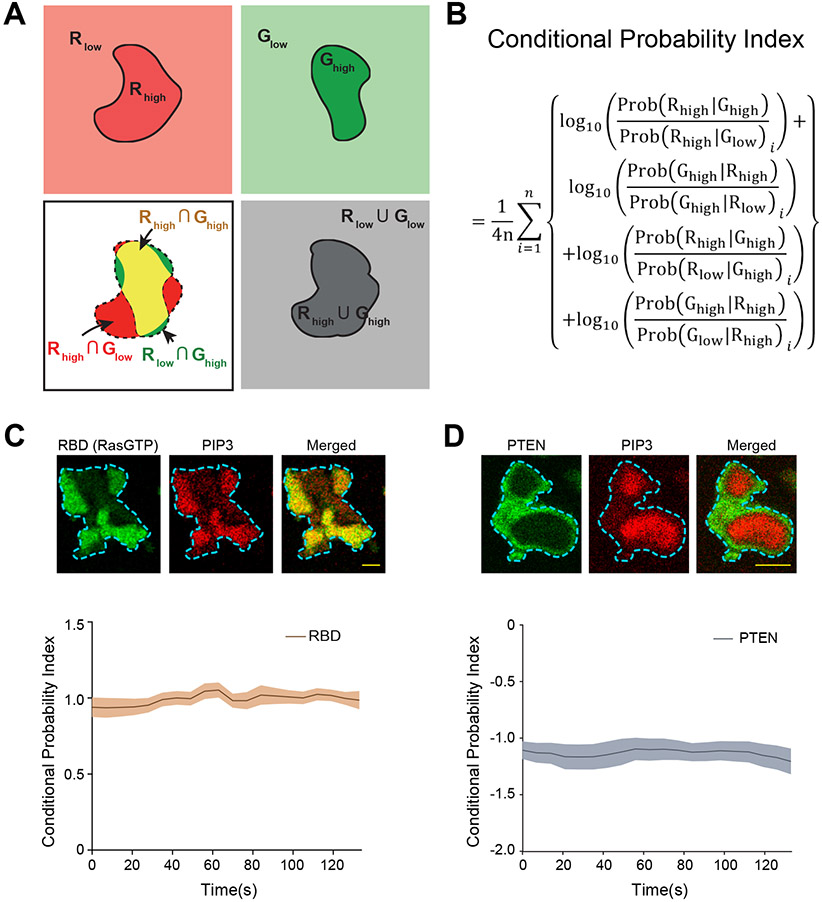
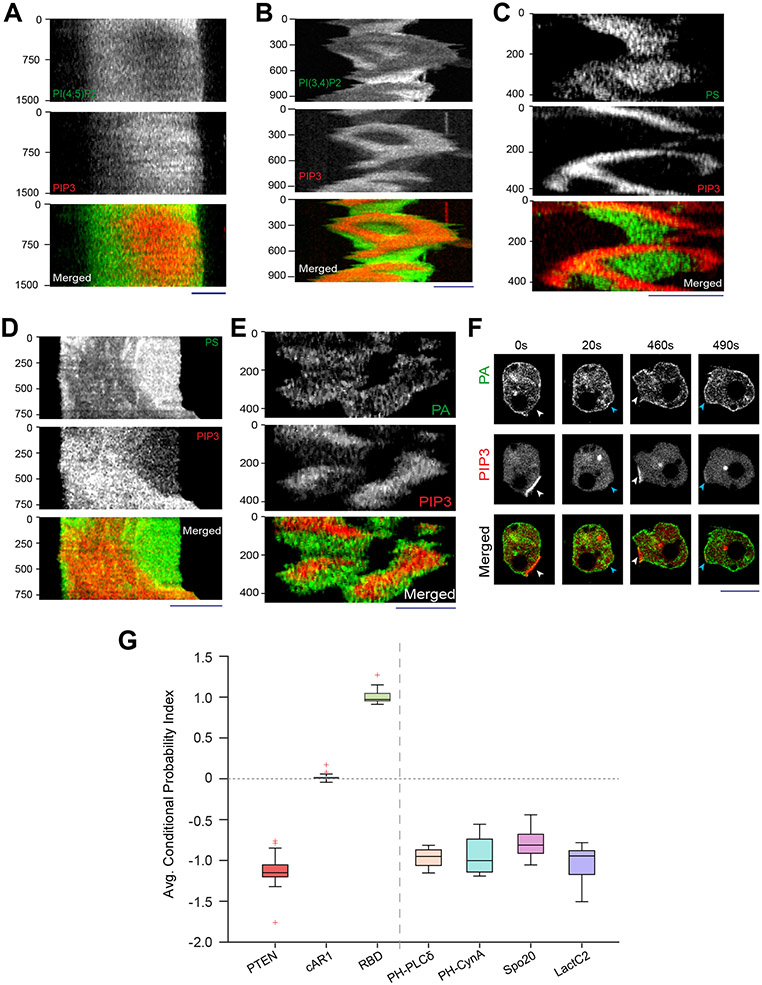
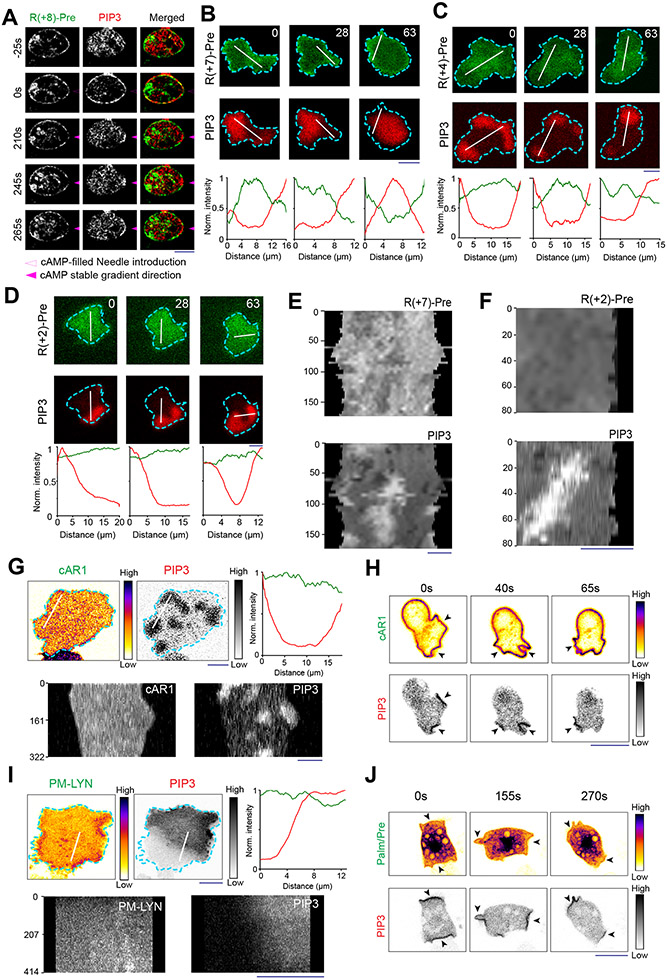
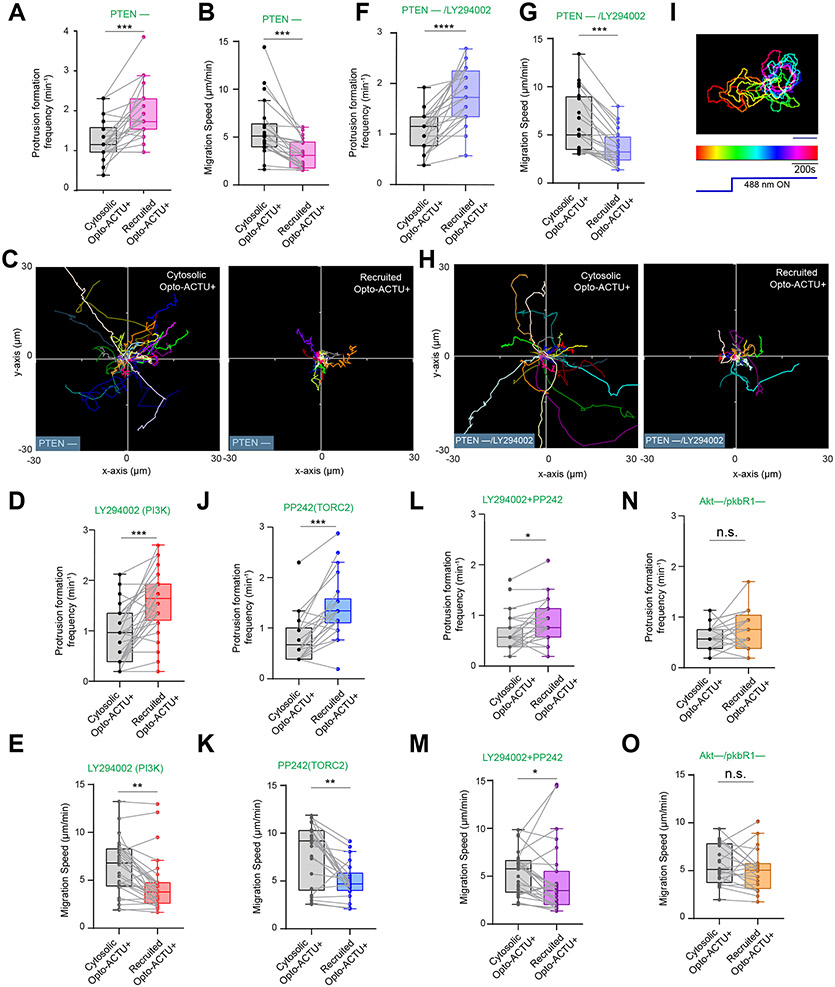
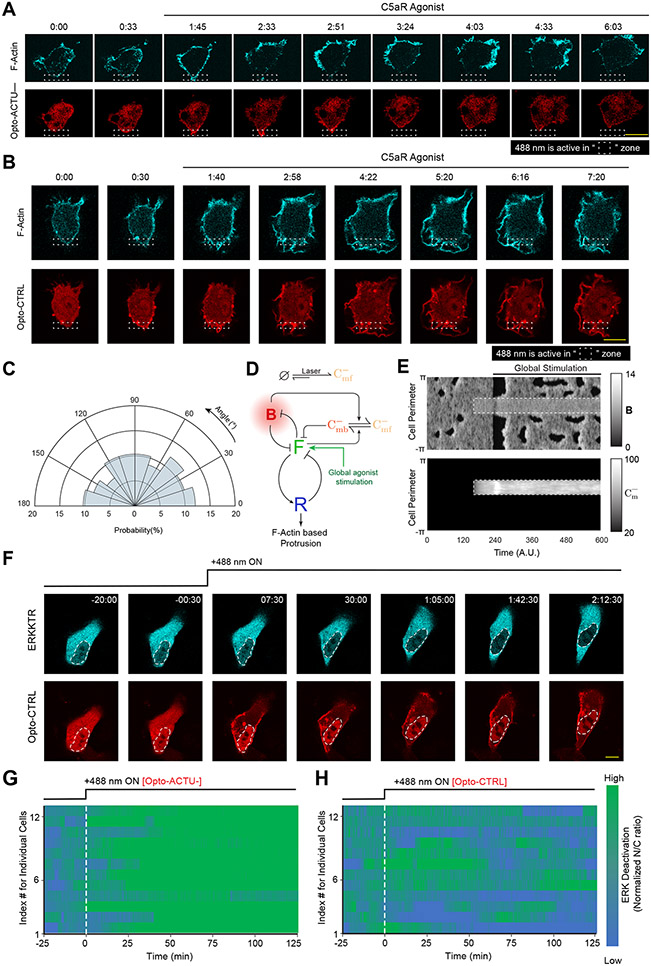
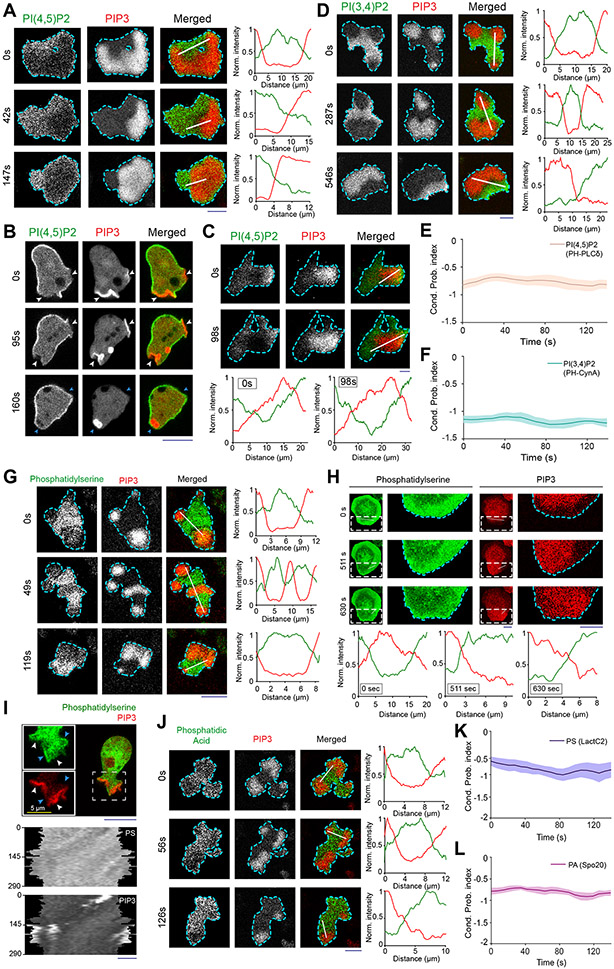
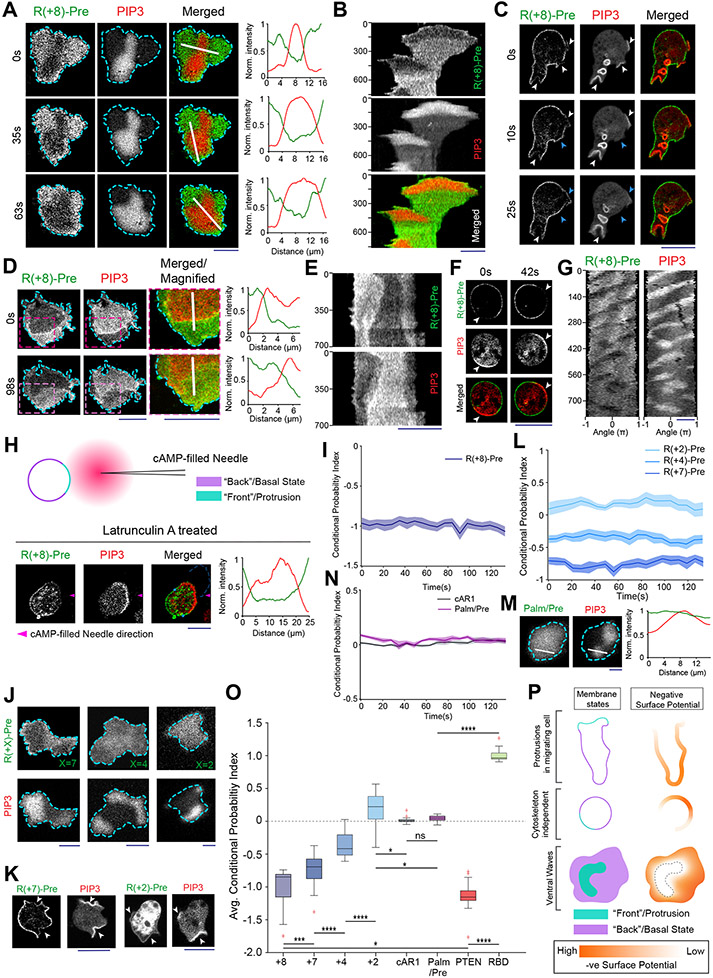
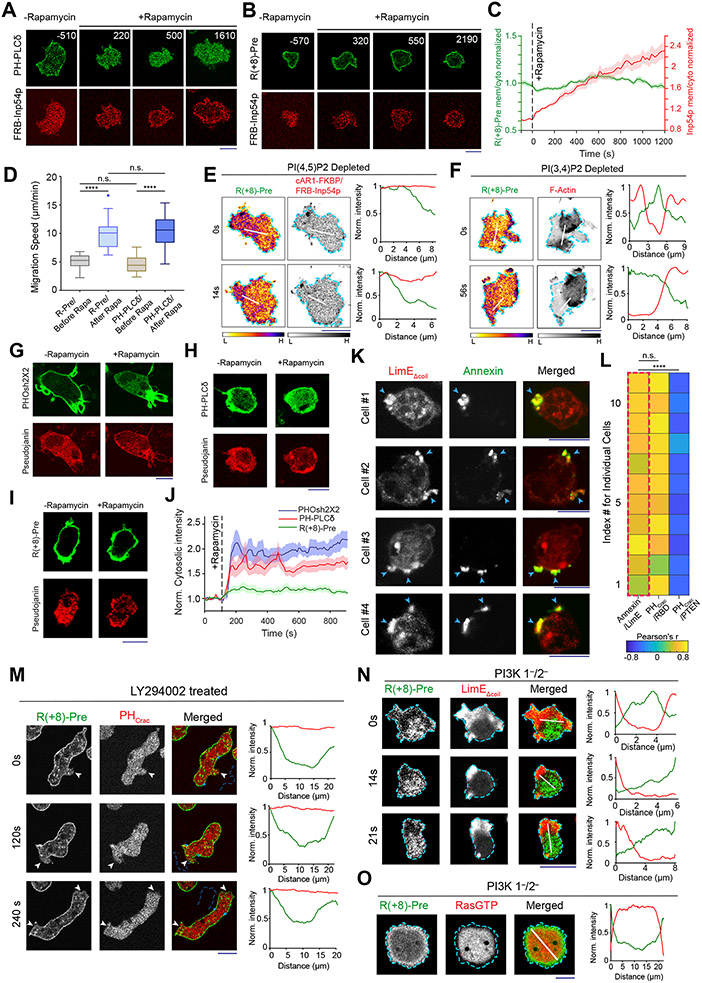
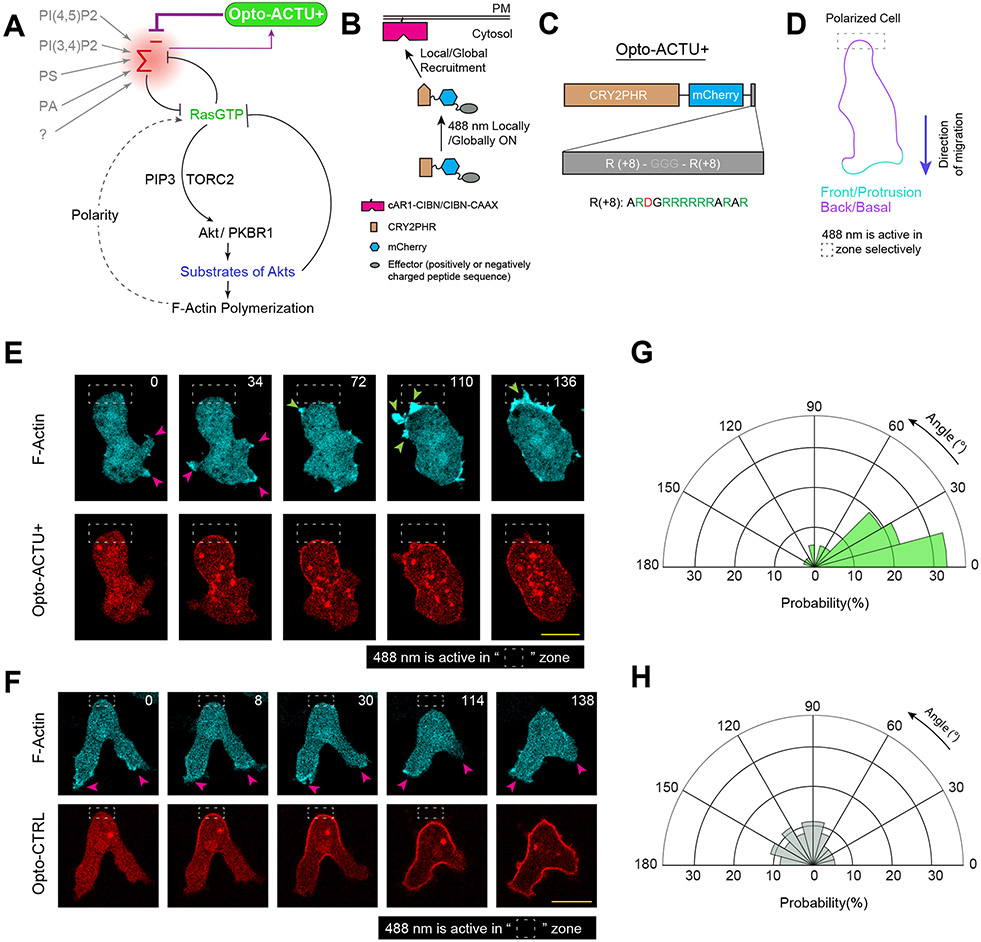
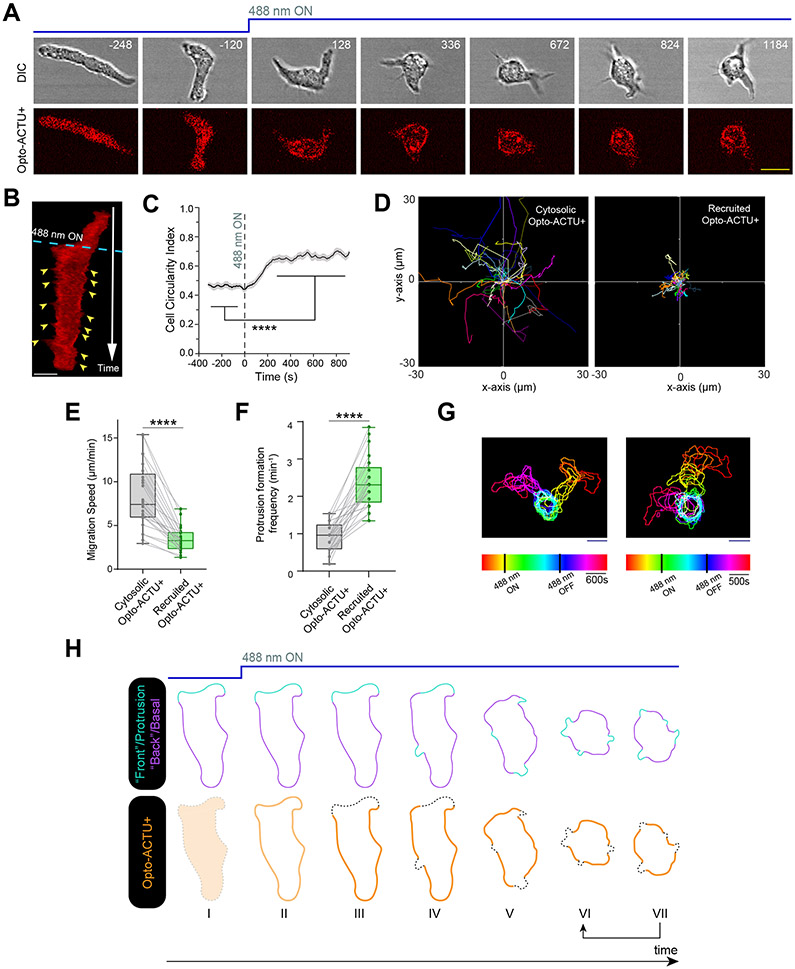
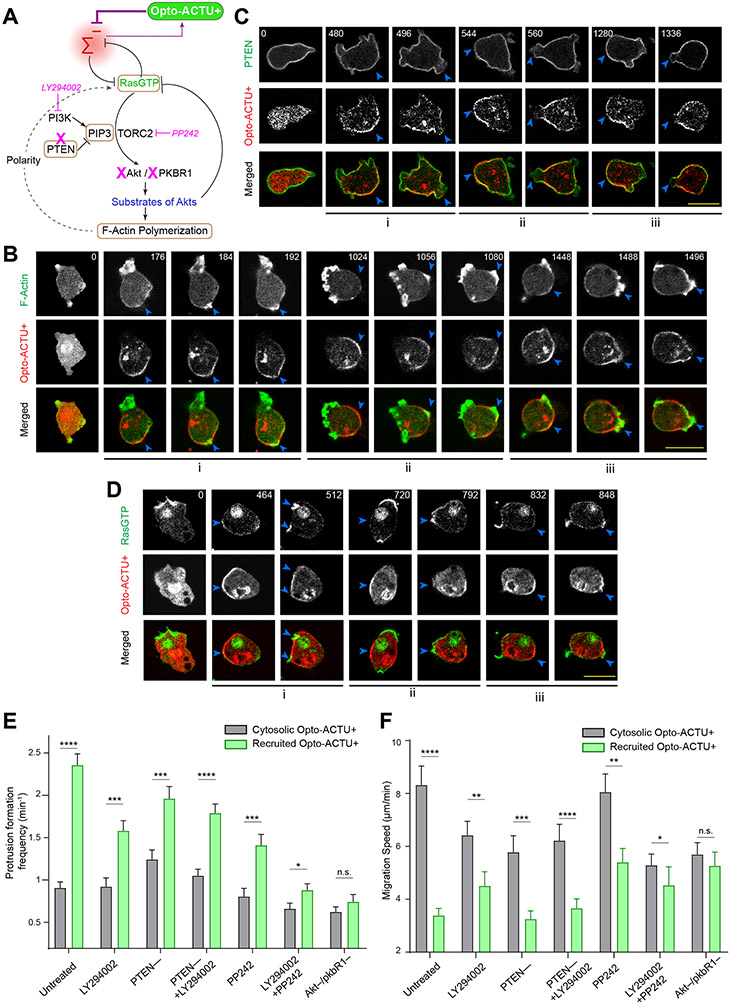
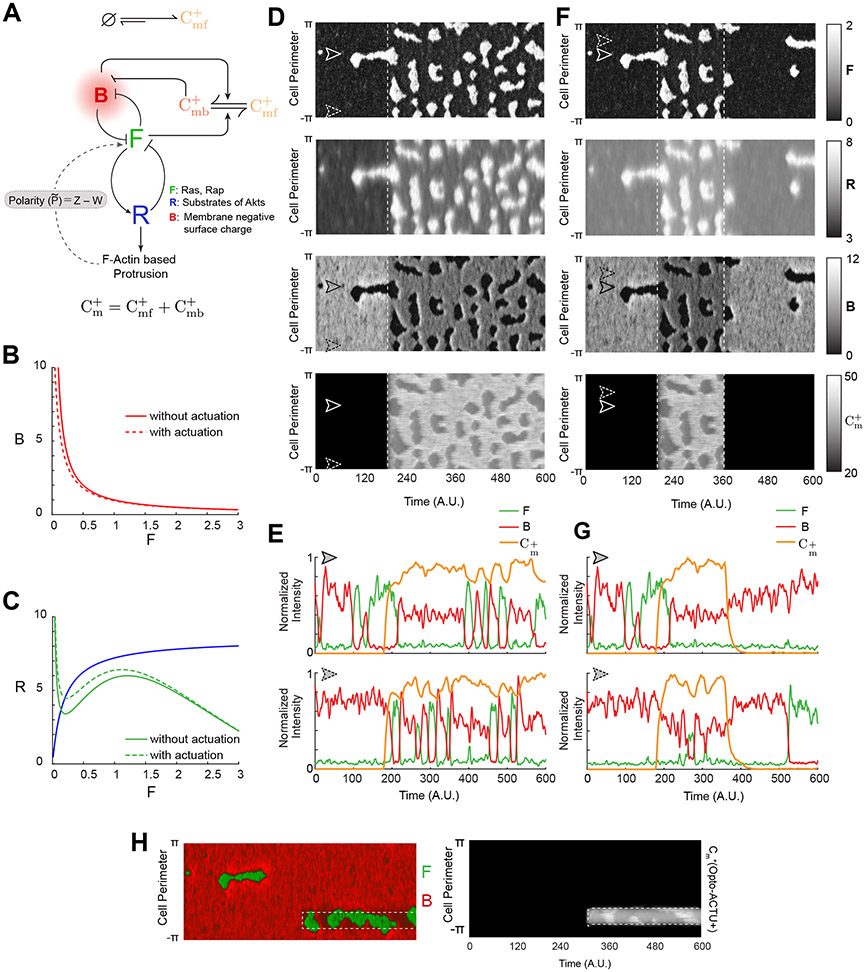
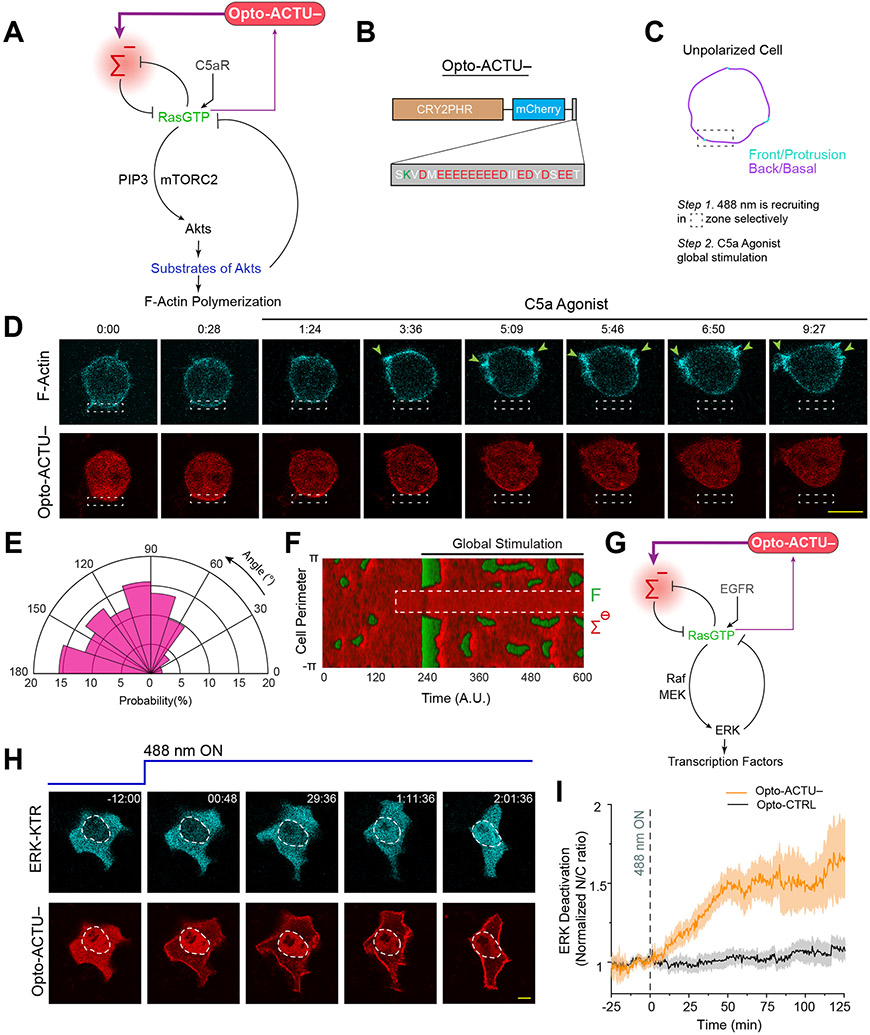
During cell migration and polarization, numerous signal transduction and cytoskeletal components self-organize to generate localized protrusions. Although biochemical and genetic analyses have delineated many specific interactions, how the activation and localization of so many different molecules are spatiotemporally orchestrated at the subcellular level has remained unclear. Here we show that the regulation of negative surface charge on the inner leaflet of the plasma membrane plays an integrative role in the molecular interactions. Surface charge, or zeta potential, is transiently lowered at new protrusions and within cortical waves of Ras/PI3K/TORC2/F-actin network activation. Rapid alterations of inner leaflet anionic phospholipids-such as PI(4,5)P2, PI(3,4)P2, phosphatidylserine and phosphatidic acid-collectively contribute to the surface charge changes. Abruptly reducing the surface charge by recruiting positively charged optogenetic actuators was sufficient to trigger the entire biochemical network, initiate de novo protrusions and abrogate pre-existing polarity. These effects were blocked by genetic or pharmacological inhibition of key signalling components such as AKT and PI3K/TORC2. Conversely, increasing the negative surface charge deactivated the network and locally suppressed chemoattractant-induced protrusions or subverted EGF-induced ERK activation. Computational simulations involving excitable biochemical networks demonstrated that slight changes in feedback loops, induced by recruitment of the charged actuators, could lead to outsized effects on system activation. We propose that key signalling network components act on, and are in turn acted upon, by surface charge, closing feedback loops, which bring about the global-scale molecular self-organization required for spontaneous protrusion formation, cell migration and polarity establishment.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Figures
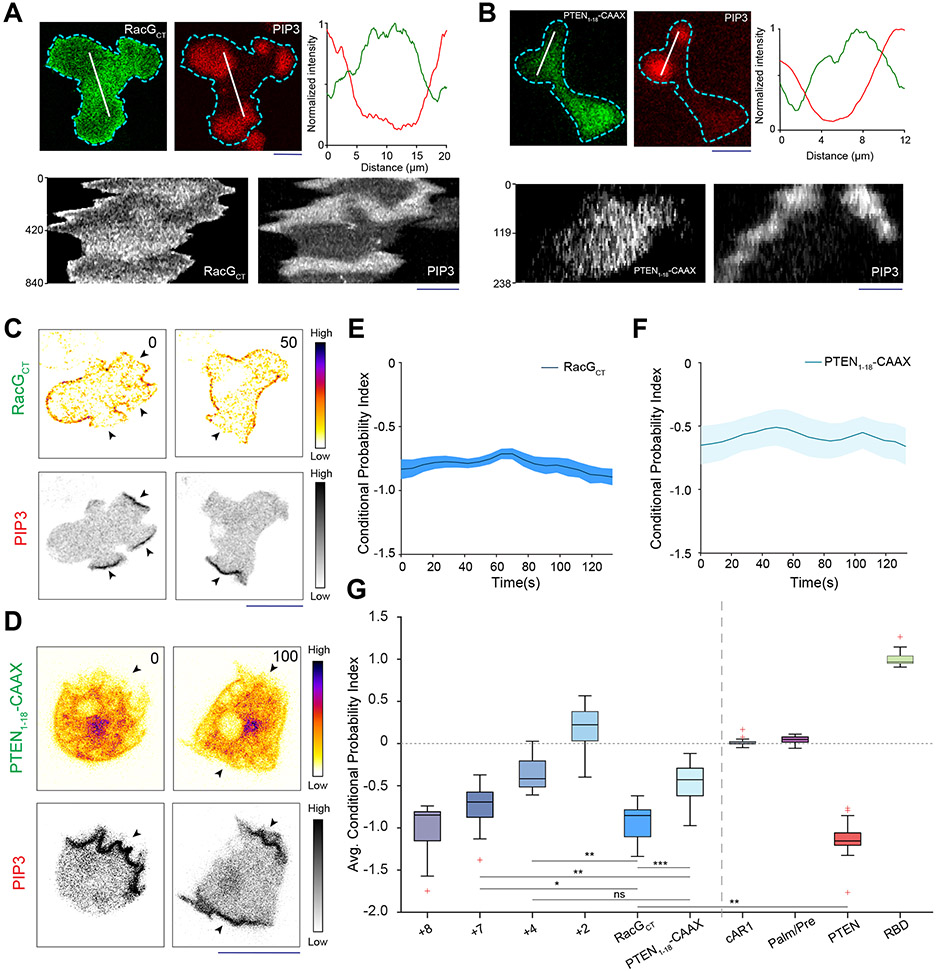
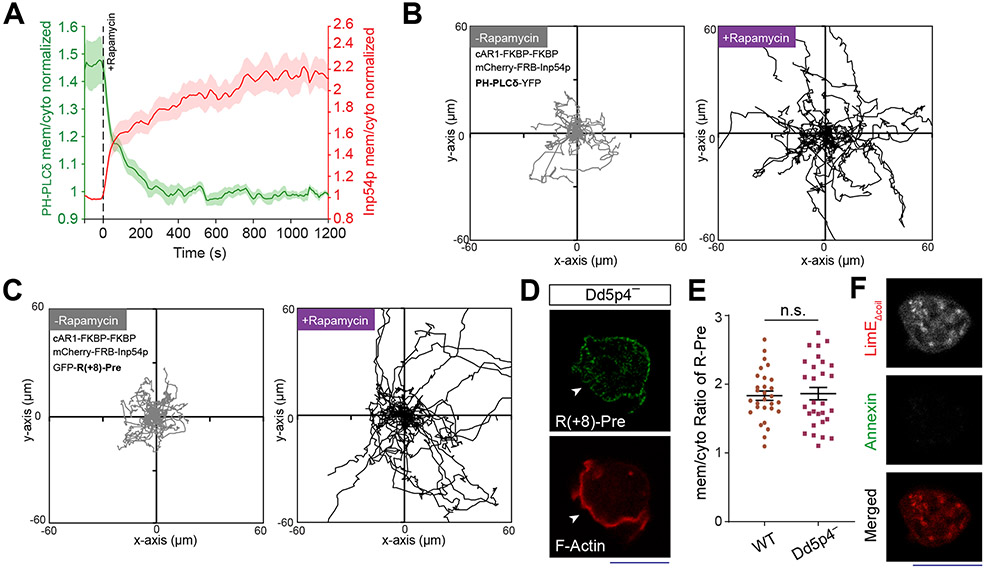
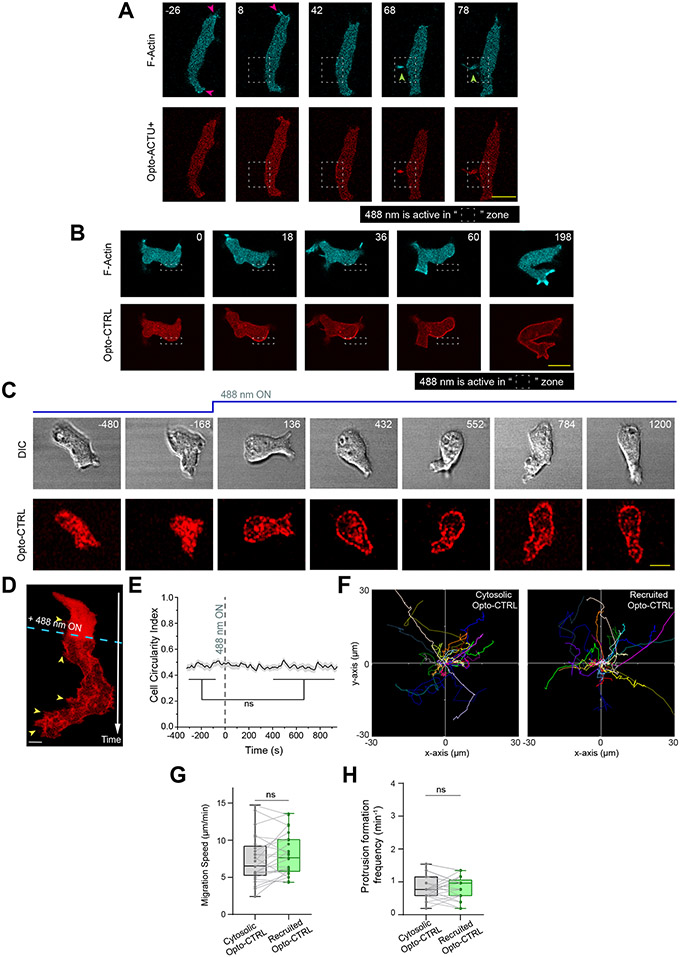
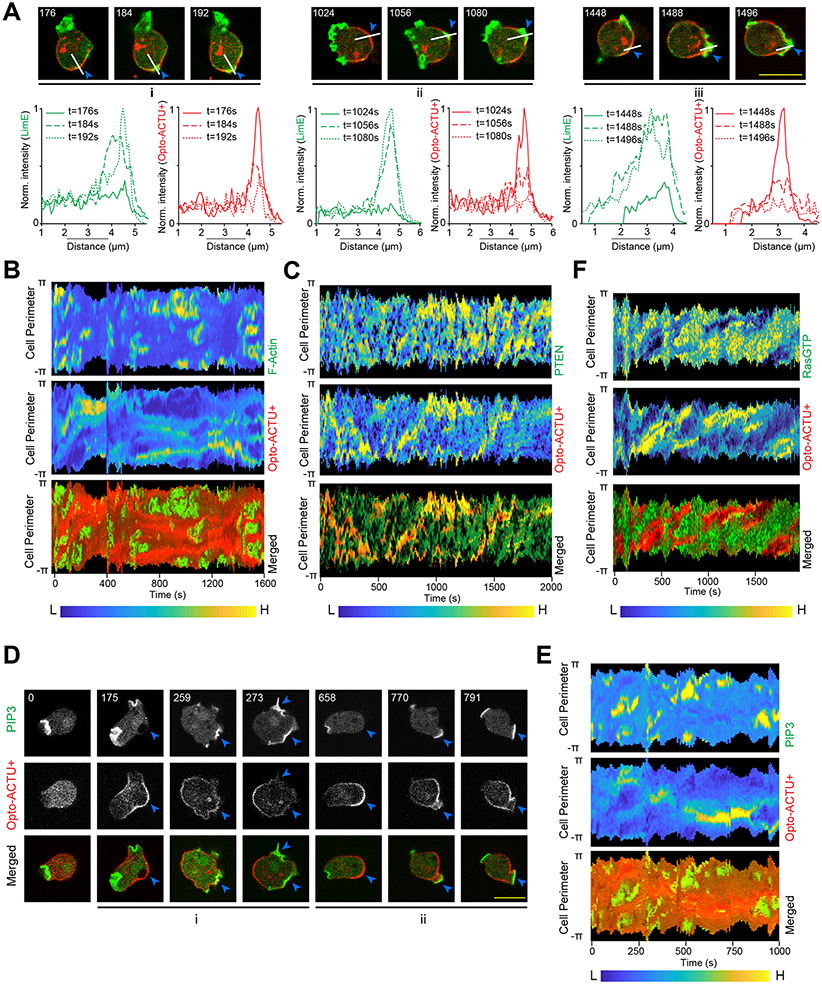
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
