

The HASTER lncRNA promoter is a cis-acting transcriptional stabilizer of HNF1A
- PMID: 36202974
- PMCID: PMC9586874
- DOI: 10.1038/s41556-022-00996-8
The HASTER lncRNA promoter is a cis-acting transcriptional stabilizer of HNF1A
Abstract
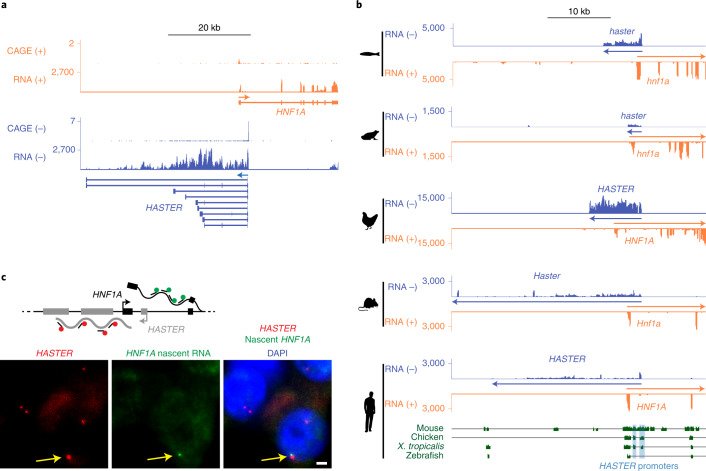
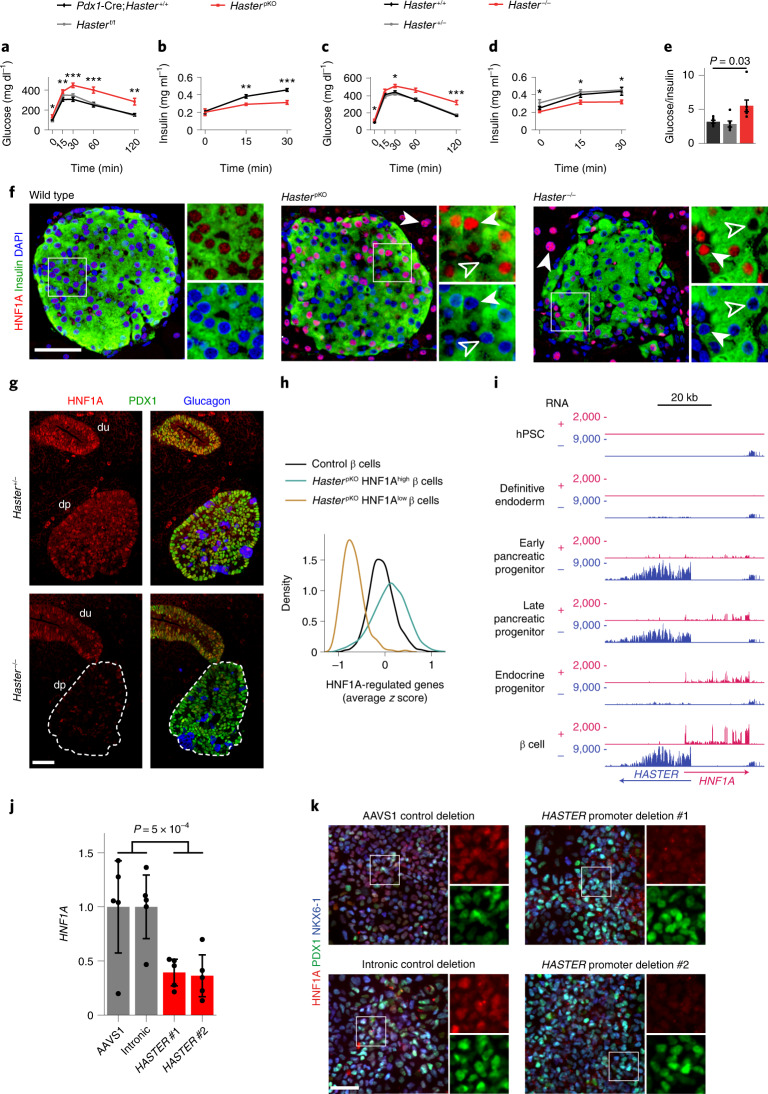
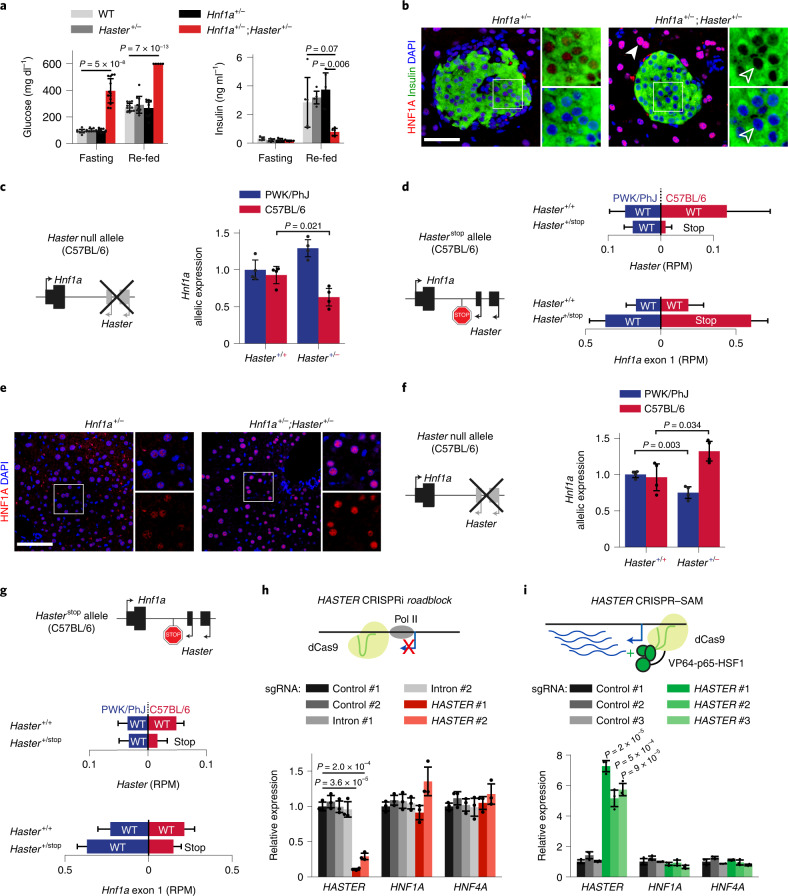
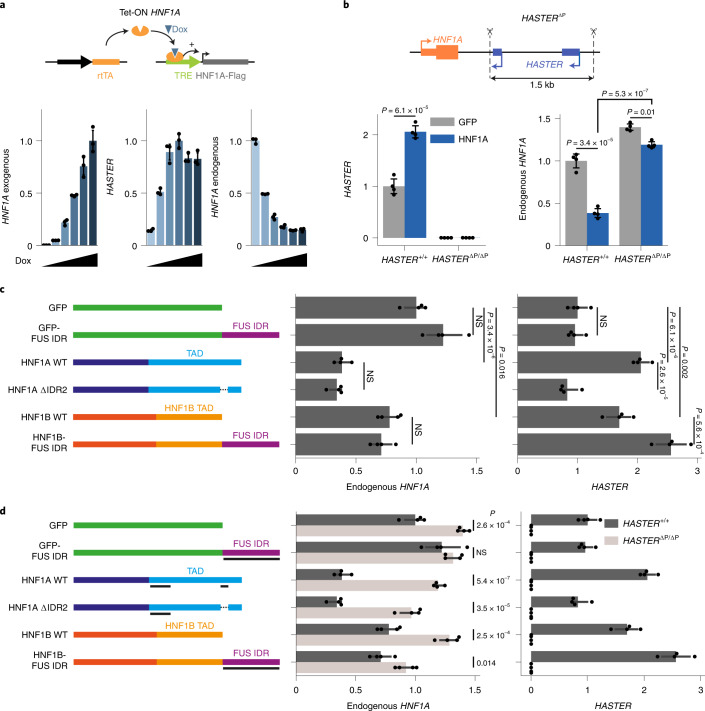
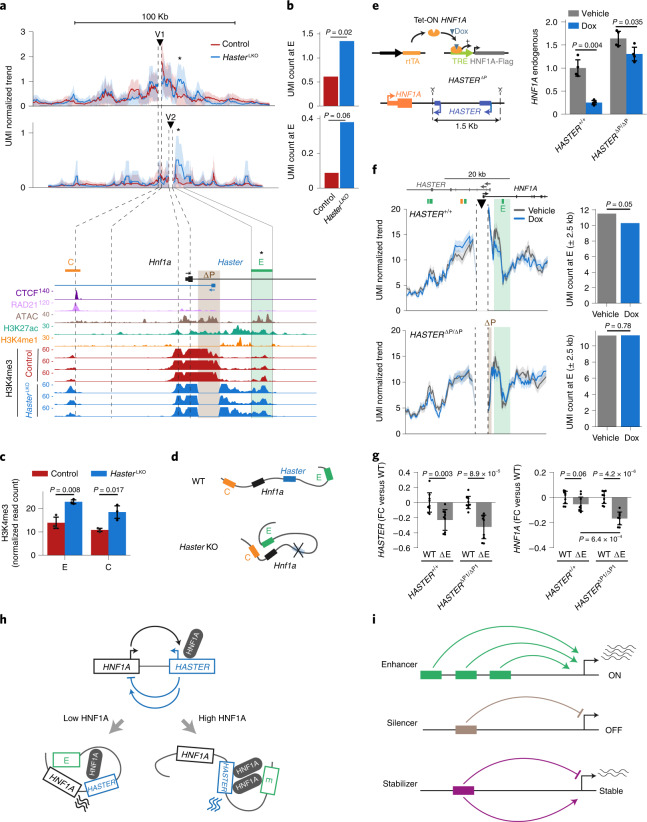
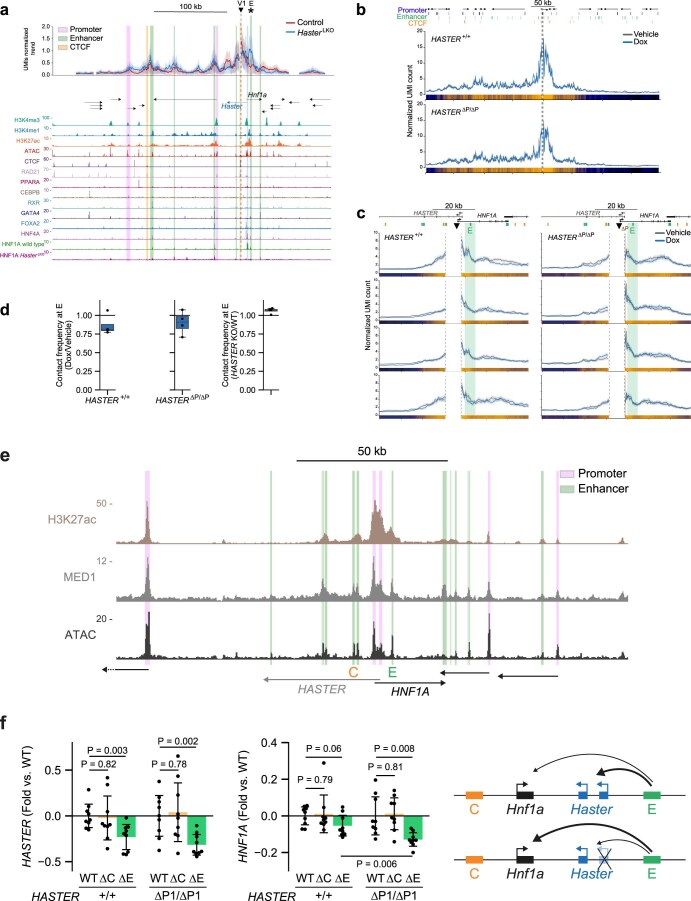
The biological purpose of long non-coding RNAs (lncRNAs) is poorly understood. Haploinsufficient mutations in HNF1A homeobox A (HNF1A), encoding a homeodomain transcription factor, cause diabetes mellitus. Here, we examine HASTER, the promoter of an lncRNA antisense to HNF1A. Using mouse and human models, we show that HASTER maintains cell-specific physiological HNF1A concentrations through positive and negative feedback loops. Pancreatic β cells from Haster mutant mice consequently showed variegated HNF1A silencing or overexpression, resulting in hyperglycaemia. HASTER-dependent negative feedback was essential to prevent HNF1A binding to inappropriate genomic regions. We demonstrate that the HASTER promoter DNA, rather than the lncRNA, modulates HNF1A promoter-enhancer interactions in cis and thereby regulates HNF1A transcription. Our studies expose a cis-regulatory element that is unlike classic enhancers or silencers, it stabilizes the transcription of its target gene and ensures the fidelity of a cell-specific transcription factor program. They also show that disruption of a mammalian lncRNA promoter can cause diabetes mellitus.
© 2022. The Author(s).
Conflict of interest statement
P.R. is a founder and consultant for EndoCells/UniverCell Biosolutions. All of the remaining authors declare no competing interests.
Figures











References
-
- Gil N, Ulitsky I. Regulation of gene expression by cis-acting long non-coding RNAs. Nat. Rev. Genet. 2020;21:102–117. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
