

Precise DNA cleavage using CRISPR-SpRYgests
- PMID: 36203014
- PMCID: PMC10023266
- DOI: 10.1038/s41587-022-01492-y
Precise DNA cleavage using CRISPR-SpRYgests
Abstract
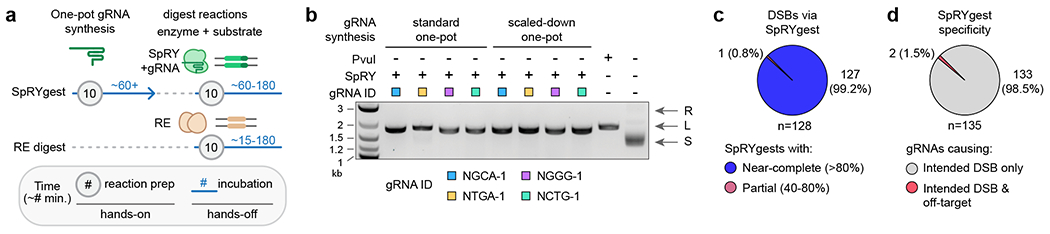
Methods for in vitro DNA cleavage and molecular cloning remain unable to precisely cleave DNA directly adjacent to bases of interest. Restriction enzymes (REs) must bind specific motifs, whereas wild-type CRISPR-Cas9 or CRISPR-Cas12 nucleases require protospacer adjacent motifs (PAMs). Here we explore the utility of our previously reported near-PAMless SpCas9 variant, named SpRY, to serve as a universal DNA cleavage tool for various cloning applications. By performing SpRY DNA digests (SpRYgests) using more than 130 guide RNAs (gRNAs) sampling a wide diversity of PAMs, we discovered that SpRY is PAMless in vitro and can cleave DNA at practically any sequence, including sites refractory to cleavage with wild-type SpCas9. We illustrate the versatility and effectiveness of SpRYgests to improve the precision of several cloning workflows, including those not possible with REs or canonical CRISPR nucleases. We also optimize a rapid and simple one-pot gRNA synthesis protocol to streamline SpRYgest implementation. Together, SpRYgests can improve various DNA engineering applications that benefit from precise DNA breaks.
© 2022. The Author(s), under exclusive licence to Springer Nature America, Inc.
Conflict of interest statement
Competing Interests Statement:
K.A.C., R.T.W., and B.P.K are inventors on patents and/or patent applications filed by Mass General Brigham that describe genome engineering technologies, including for the development of SpRY (R.T.W. and B.P.K.). B.P.K. is a consultant for EcoR1 capital and is an advisor to Acrigen Biosciences, Life Edit Therapeutics, and Prime Medicine. L.P. has financial interests in Edilytics and SeQure Dx, Inc. L.P.’s interests were reviewed and are managed by Massachusetts General Hospital and Partners HealthCare in accordance with their conflict-of-interest policies. M.M. and G.B.R. are employees of the Research Department at New England Biolabs Inc. (NEB). NEB is a commercial supplier of molecular biology reagents including some that have been used in this work. The remaining authors declare no competing interests.
Figures



References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
