

Transcriptome profiling of Arabidopsis slac1-3 mutant reveals compensatory alterations in gene expression underlying defective stomatal closure
- PMID: 36204078
- PMCID: PMC9530288
- DOI: 10.3389/fpls.2022.987606
Transcriptome profiling of Arabidopsis slac1-3 mutant reveals compensatory alterations in gene expression underlying defective stomatal closure
Abstract
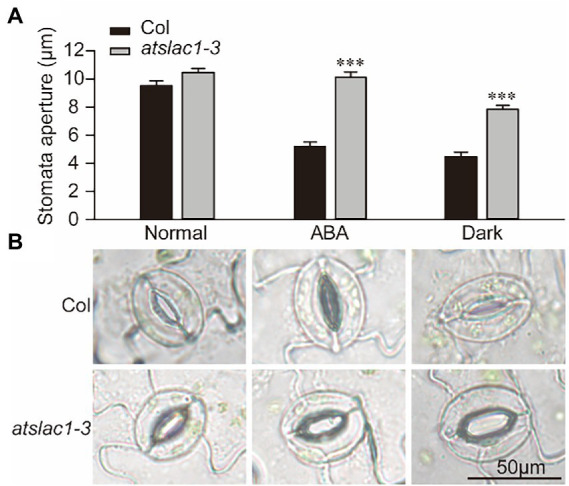
Plants adjust their stomatal aperture for regulating CO2 uptake and transpiration. S-type anion channel SLAC1 (slow anion channel-associated 1) is required for stomatal closure in response to various stimuli such as abscisic acid, CO2, and light/dark transitions etc. Arabidopsis slac1 mutants exhibited defects in stimulus-induced stomatal closure, reduced sensitivity to darkness, and faster water loss from detached leaves. The global transcriptomic response of a plant with defective stimuli-induced stomatal closure (particularly because of defects in SLAC1) remains to be explored. In the current research we attempted to address the same biological question by comparing the global transcriptomic changes in Arabidopsis slac1-3 mutant and wild-type (WT) under dark, and dehydration stress, using RNA-sequencing. Abscisic acid (ABA)- and dark-induced stomatal closure was defective in Arabidopsis slac1-3 mutants, consequently the mutants had cooler leaf temperature than WT. Next, we determined the transcriptomic response of the slac1-3 mutant and WT under dark and dehydration stress. Under dehydration stress, the molecular response of slac1-3 mutant was clearly distinct from WT; the number of differentially expressed genes (DEGs) was significantly higher in mutant than WT. Dehydration induced DEGs in mutant were related to hormone signaling pathways, and biotic and abiotic stress response. Although, overall number of DEGs in both genotypes was not different under dark, however, the expression pattern was very much distinct; whereas majority of DEGs in WT were found to be downregulated, in slac1-3 majority were upregulated under dark. Further, a set 262 DEGs was identified with opposite expression pattern between WT and mutant under light-darkness transition. Amongst these, DEGs belonging to stress hormone pathways, and biotic and abiotic stress response were over-represented. To sum up, we have reported gene expression reprogramming underlying slac1-3 mutation and resultantly defective stomatal closure in Arabidopsis. Moreover, the induction of biotic and abiotic response in mutant under dehydration and darkness could be suggestive of the role of stomata as a switch in triggering these responses. To summarize, the data presented here provides useful insights into the gene expression reprogramming underlying slac1-3 mutation and resultant defects in stomatal closure.
Keywords: abscisic acid; anion channel; drought stress; stomata; transcriptome.
Copyright © 2022 Wang, Ouyang, Ren, Wang, Xu, Xin, Hussain and Qi.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
-
- Brandt B., Brodsky D. E., Xue S., Negi J., Iba K., Kangasjärvi J., et al. . (2012). Reconstitution of abscisic acid activation of SLAC1 anion channel by CPK6 and OST1 kinases and branched ABI1 PP2C phosphatase action. Proc. Natl. Acad. Sci. U. S. A. 109, 10593–10598. doi: 10.1073/pnas.1116590109, PMID: - DOI - PMC - PubMed
-
- Cheong Y. H., Pandey G. K., Grant J. J., Batistic O., Li L., Kim B. G., et al. . (2007). Two calcineurin B-like calcium sensors, interacting with protein kinase CIPK23, regulate leaf transpiration and root potassium uptake in Arabidopsis. Plant J. 52, 223–239. doi: 10.1111/j.1365-313X.2007.03236.x, PMID: - DOI - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases