

Intracellular mRNA phase separation induced by cationic polymers for tumor immunotherapy
- PMID: 36209170
- PMCID: PMC9548170
- DOI: 10.1186/s12951-022-01647-8
Intracellular mRNA phase separation induced by cationic polymers for tumor immunotherapy
Abstract
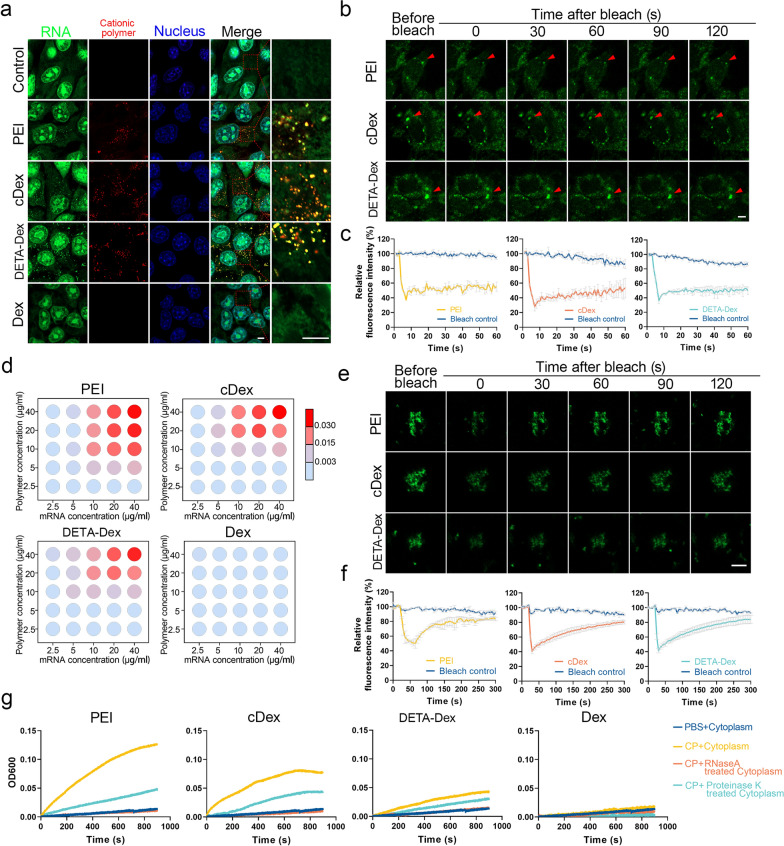
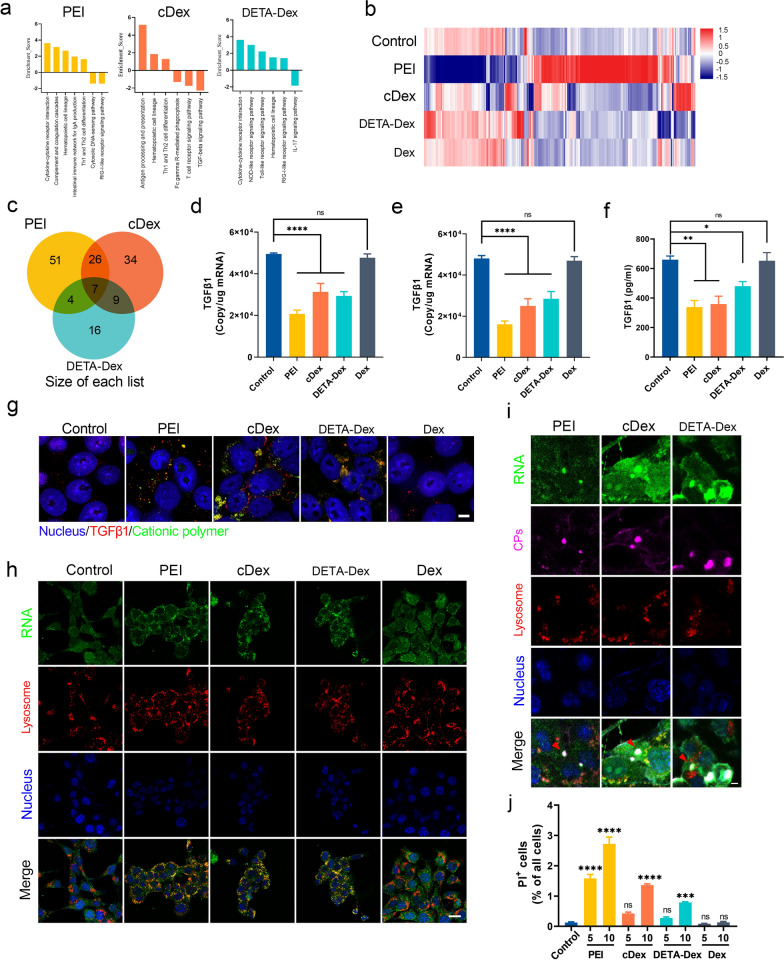
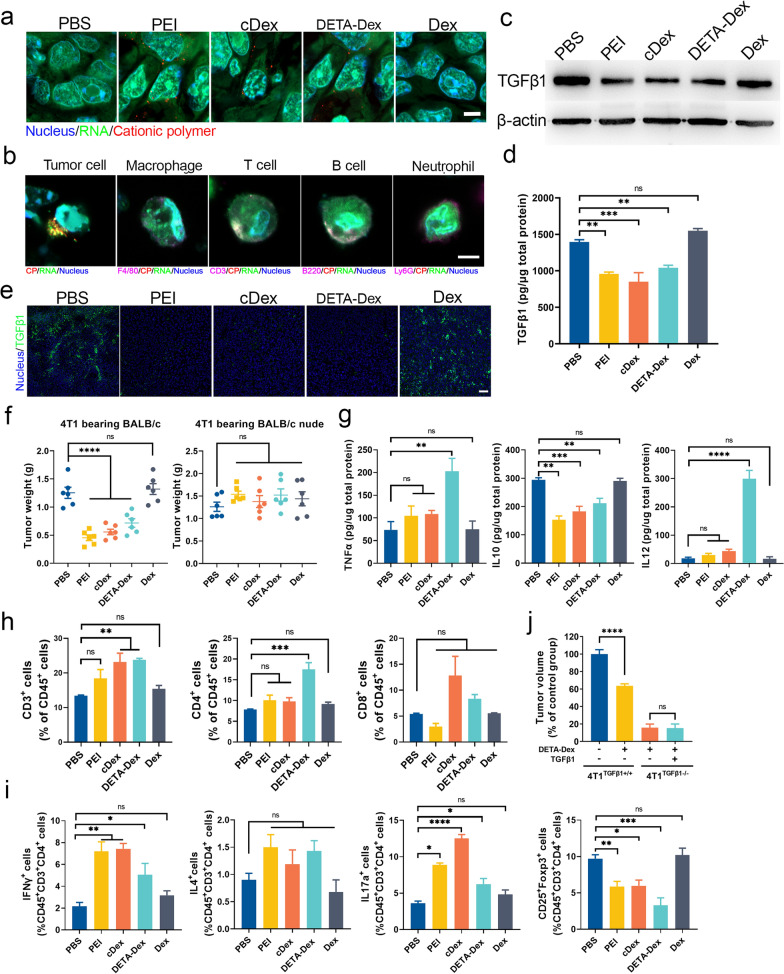
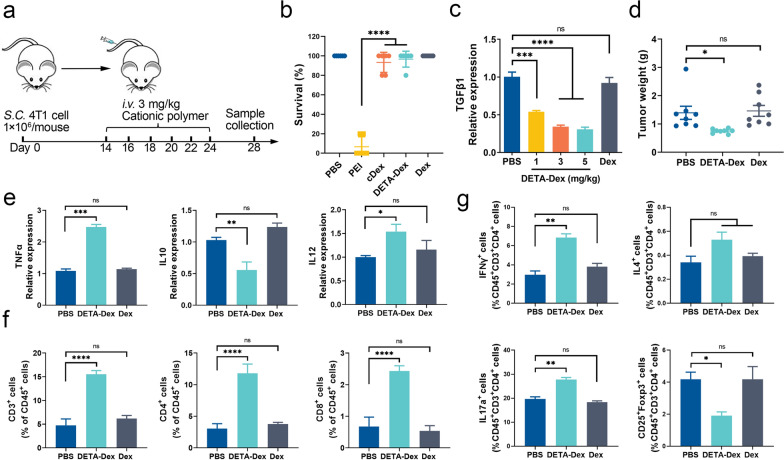
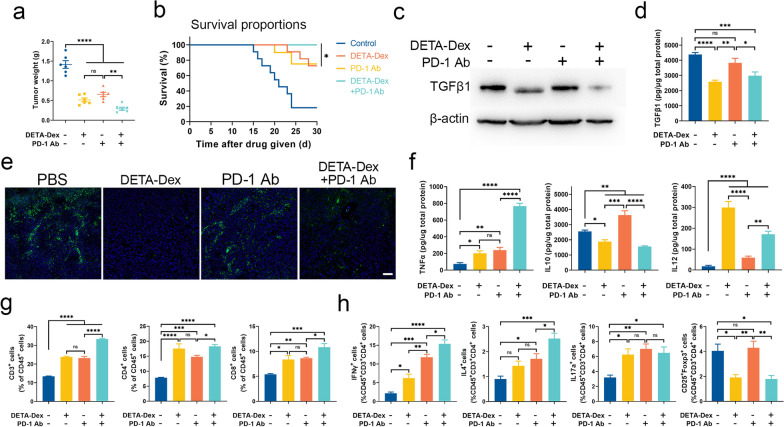
The formation of biomolecular condensates via liquid‒liquid phase separation (LLPS) is an advantageous strategy for cells to organize their subcellular compartments for diverse functions. Recent findings suggest that RNA or RNA-related LLPS techniques have potential for the development of new cellular regulation strategies. However, manipulating RNA LLPS in living cells has great challenges. Herein, we report that cationic polymers (CPs) have strong RNA LLPS-inducing activity. By introducing CPs into living cells or RNA solutions, significant RNA LLPS was verified through confocal imaging, turbidity assays, and fluorescence recovery after photobleaching (FRAP) tests. Among them, turbidity kinetics determinations indicated that the hydrophilic positively charged amino groups on the CPs play essential roles in RNA phase separation. Moreover, the LLPS induced by the cationic polymers dramatically changed the gene expression patterns in the cells. Interestingly, we found that TGFβ1 mRNA was highly encapsulated in the RNA droplets, which lowered the immunosuppressive capability of the tumor cells and triggered marked antitumor reactions in a mouse breast cancer model. Thus, we present here the CP-based modulation of RNA LLPS as a novel transcriptional manipulation method with potential for cancer immunotherapy drug development.
Keywords: Cationic polymers; Immunotherapy; TGFβ1; Tumor microenvironment; mRNA phase separation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
N6-methyladenosine RNA methylation modulates liquid‒liquid phase separation in plants.Plant Cell. 2023 Sep 1;35(9):3205-3213. doi: 10.1093/plcell/koad103. Plant Cell. 2023. PMID: 37032432 Free PMC article. Review.
-
How do RNA binding proteins trigger liquid-liquid phase separation in human health and diseases?Biosci Trends. 2022 Dec 26;16(6):389-404. doi: 10.5582/bst.2022.01449. Epub 2022 Dec 3. Biosci Trends. 2022. PMID: 36464283 Review.
-
Liquid-liquid phase separation in plants: Advances and perspectives from model species to crops.Plant Commun. 2024 Jan 8;5(1):100663. doi: 10.1016/j.xplc.2023.100663. Epub 2023 Jul 26. Plant Commun. 2024. PMID: 37496271 Free PMC article. Review.
-
Fixation can change the appearance of phase separation in living cells.Elife. 2022 Nov 29;11:e79903. doi: 10.7554/eLife.79903. Elife. 2022. PMID: 36444977 Free PMC article.
-
[Latest Findings on Phase Separation of Cytomechanical Proteins].Sichuan Da Xue Xue Bao Yi Xue Ban. 2024 Jan 20;55(1):19-23. doi: 10.12182/20240160206. Sichuan Da Xue Xue Bao Yi Xue Ban. 2024. PMID: 38322526 Free PMC article. Chinese.
Cited by
-
The Burgeoning Significance of Liquid-Liquid Phase Separation in the Pathogenesis and Therapeutics of Cancers.Int J Biol Sci. 2024 Feb 12;20(5):1652-1668. doi: 10.7150/ijbs.92988. eCollection 2024. Int J Biol Sci. 2024. PMID: 38481812 Free PMC article. Review.
-
Systemic Delivery of Paclitaxel by Find-Me Nanoparticles Activates Antitumor Immunity and Eliminates Tumors.ACS Nano. 2024 Jan 30;18(4):3681-3698. doi: 10.1021/acsnano.3c11445. Epub 2024 Jan 16. ACS Nano. 2024. PMID: 38227965 Free PMC article.
-
Liquid-liquid phase separation: an emerging perspective on the tumorigenesis, progression, and treatment of tumors.Front Immunol. 2025 Jun 26;16:1604015. doi: 10.3389/fimmu.2025.1604015. eCollection 2025. Front Immunol. 2025. PMID: 40642070 Free PMC article. Review.
-
Role of liquid-liquid phase separation in cancer: Mechanisms and therapeutic implications.Cancer Innov. 2024 Sep 17;3(5):e144. doi: 10.1002/cai2.144. eCollection 2024 Oct. Cancer Innov. 2024. PMID: 39290787 Free PMC article. Review.
-
RNA: The Unsuspected Conductor in the Orchestra of Macromolecular Crowding.Chem Rev. 2024 Apr 24;124(8):4734-4777. doi: 10.1021/acs.chemrev.3c00575. Epub 2024 Apr 5. Chem Rev. 2024. PMID: 38579177 Free PMC article. Review.
References
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
