

Cross-species evolution of a highly potent AAV variant for therapeutic gene transfer and genome editing
- PMID: 36210364
- PMCID: PMC9548504
- DOI: 10.1038/s41467-022-33745-4
Cross-species evolution of a highly potent AAV variant for therapeutic gene transfer and genome editing
Abstract
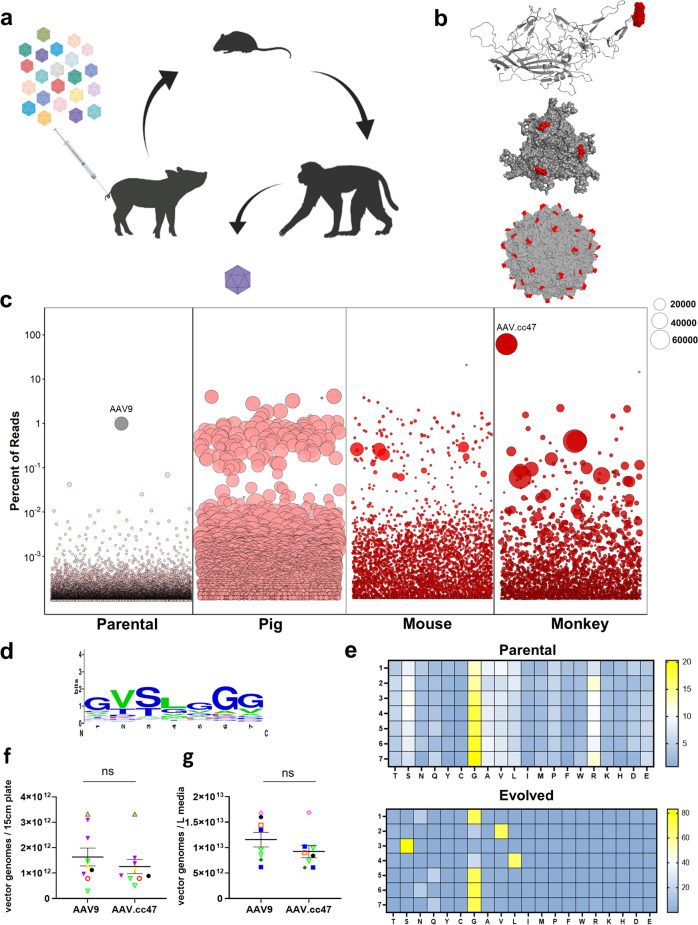
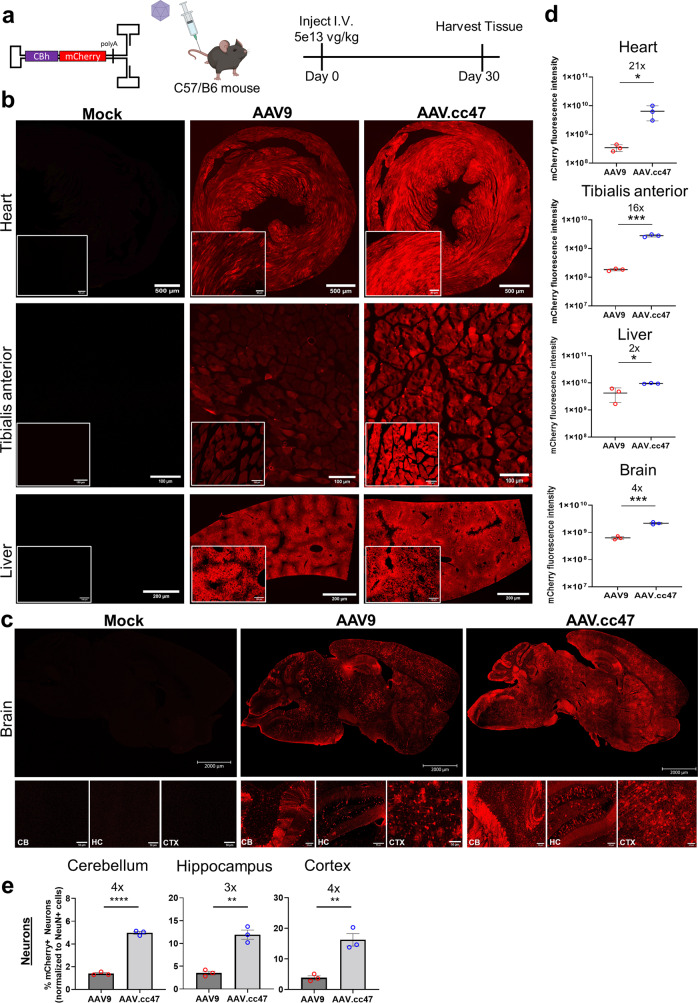
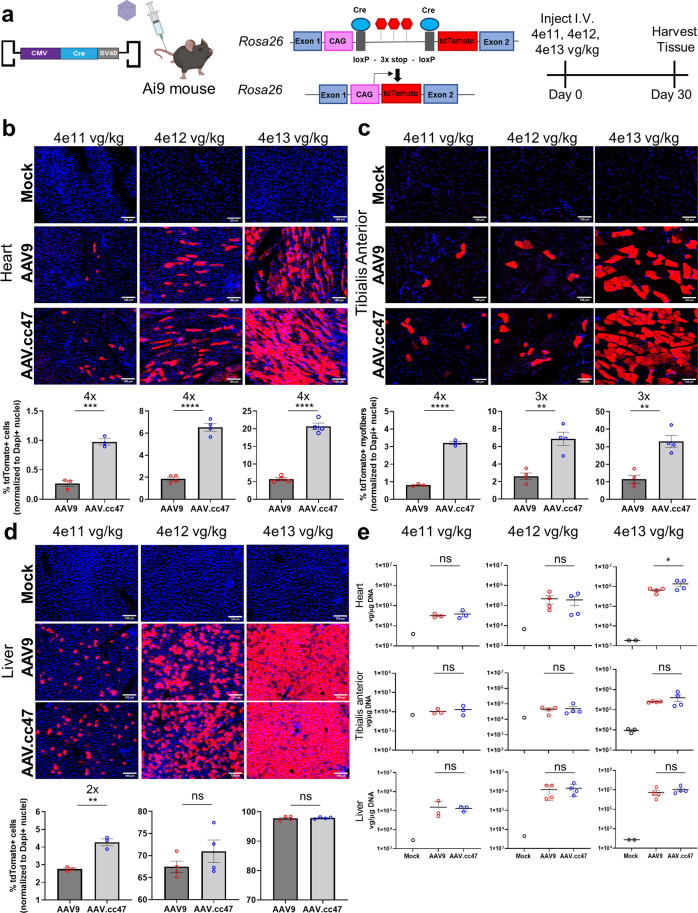
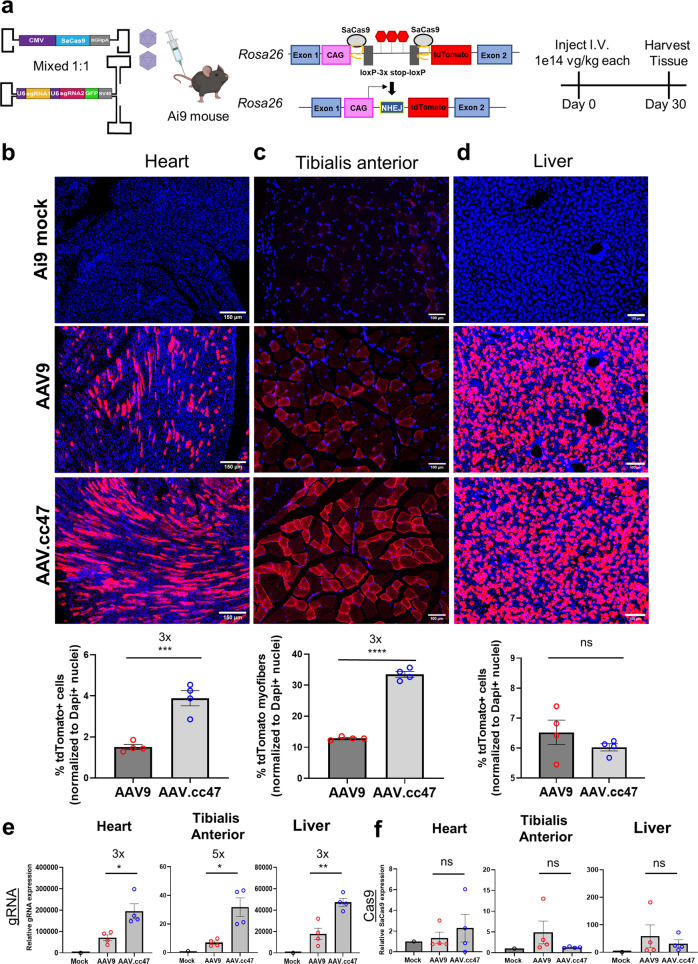
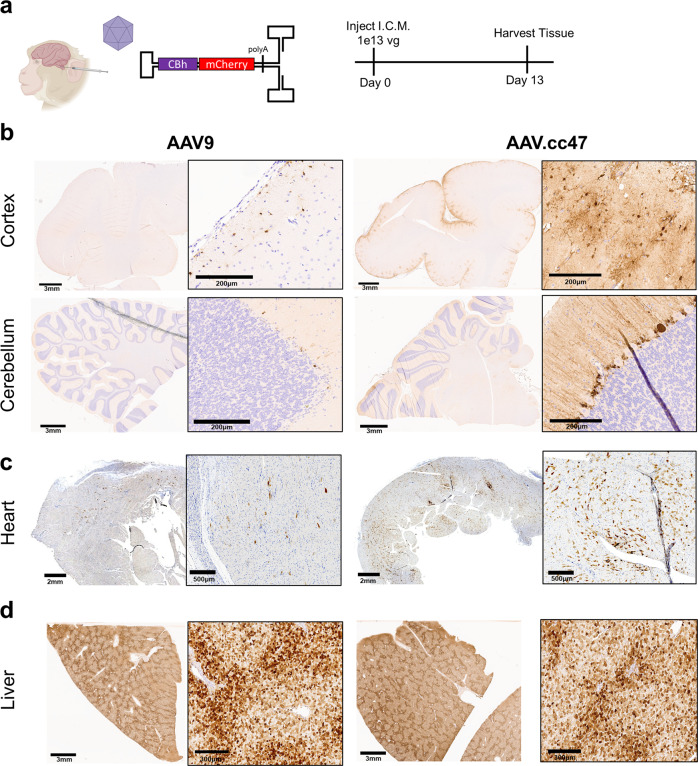
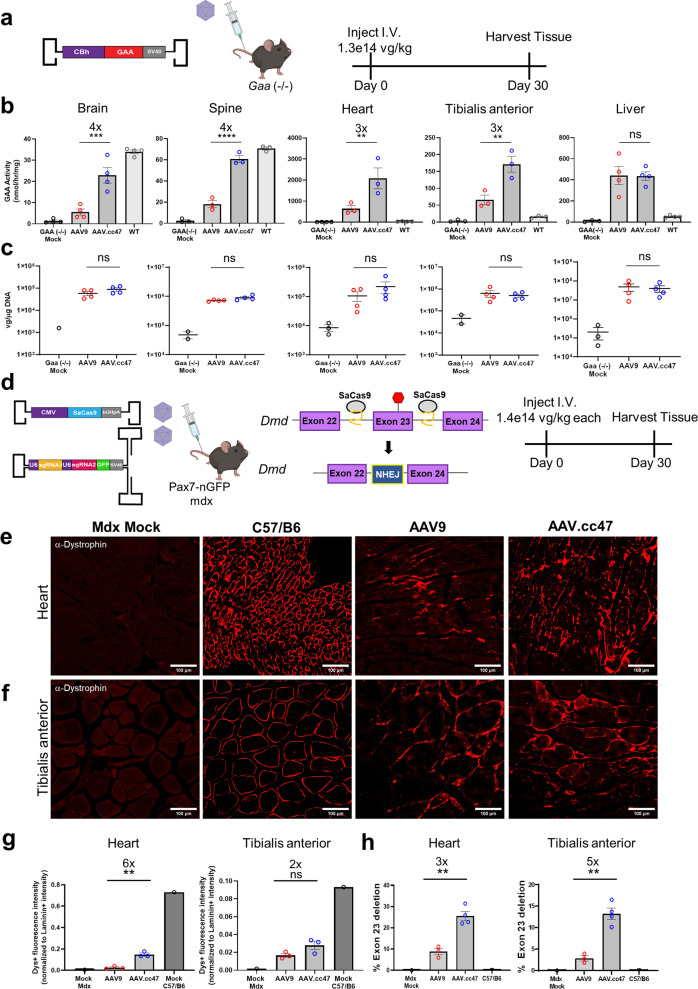
Recombinant adeno-associated viral (AAV) vectors are a promising gene delivery platform, but ongoing clinical trials continue to highlight a relatively narrow therapeutic window. Effective clinical translation is confounded, at least in part, by differences in AAV biology across animal species. Here, we tackle this challenge by sequentially evolving AAV capsid libraries in mice, pigs and macaques. We discover a highly potent, cross-species compatible variant (AAV.cc47) that shows improved attributes benchmarked against AAV serotype 9 as evidenced by robust reporter and therapeutic gene expression, Cre recombination and CRISPR genome editing in normal and diseased mouse models. Enhanced transduction efficiency of AAV.cc47 vectors is further corroborated in macaques and pigs, providing a strong rationale for potential clinical translation into human gene therapies. We envision that ccAAV vectors may not only improve predictive modeling in preclinical studies, but also clinical translatability by broadening the therapeutic window of AAV based gene therapies.
© 2022. The Author(s).
Conflict of interest statement
T.J.G, L.P.H., and A.A. have filed patent applications on the subject matter of this manuscript. A.A. is a co-founder at StrideBio and TorqueBio and serves on the advisory boards of Atsena Therapeutics, Isolere Bio, Mammoth Bio, Ring Therapeutics, and Kriya Therapeutics. C.A.G. is an advisor to Sarepta Therapeutics, Tune Therapeutics, Levo Therapeutics, and Iveric Bio and a co-founder of Tune Therapeutics, Element Genomics, and Locus Biosciences. R.M.C.R. and M.S.M. were employed at StideBio Inc., at this time. The remaining authors declare no competing interests.
Figures
References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
