

Progression in Time of Dentate Gyrus Granule Cell Layer Widening due to Excitotoxicity Occurs along In Vivo LTP Reinstatement and Contextual Fear Memory Recovery
- PMID: 36213614
- PMCID: PMC9533134
- DOI: 10.1155/2022/7432842
Progression in Time of Dentate Gyrus Granule Cell Layer Widening due to Excitotoxicity Occurs along In Vivo LTP Reinstatement and Contextual Fear Memory Recovery
Abstract
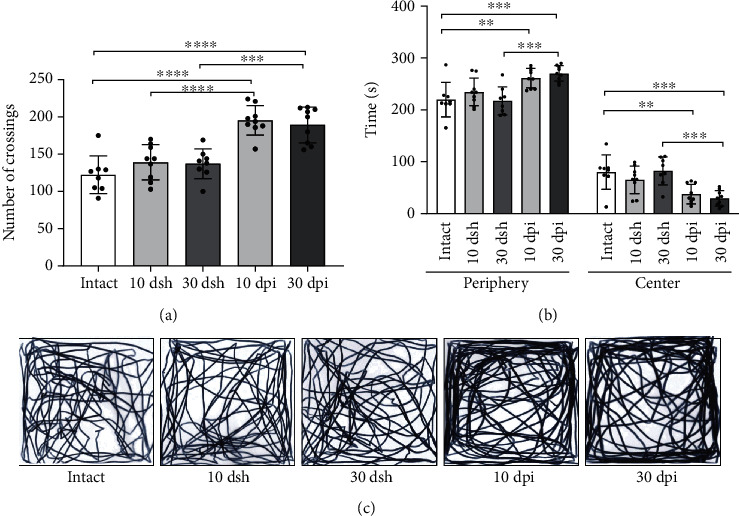
The dentate gyrus (DG) is the gateway of sensory information arriving from the perforant pathway (PP) to the hippocampus. The adequate integration of incoming information into the DG is paramount in the execution of hippocampal-dependent cognitive functions. An abnormal DG granule cell layer (GCL) widening due to granule cell dispersion has been reported under hyperexcitation conditions in animal models as well as in patients with mesial temporal lobe epilepsy, but also in patients with no apparent relation to epilepsy. Strikingly, it is unclear whether the presence and severity of GCL widening along time affect synaptic processing arising from the PP and alter the performance in hippocampal-mediated behaviors. To evaluate the above, we injected excitotoxic kainic acid (KA) unilaterally into the DG of mice and analyzed the evolution of GCL widening at 10 and 30 days post injection (dpi), while analyzing if KA-induced GCL widening affected in vivo long-term potentiation (LTP) in the PP-DG pathway, as well as the performance in learning and memory through contextual fear conditioning. Our results show that at 10 dpi, when a subtle GCL widening was observed, LTP induction, as well as contextual fear memory, were impaired. However, at 30 dpi when a pronounced increase in GCL widening was found, LTP induction and contextual fear memory were already reestablished. These results highlight the plastic potential of the DG to recover some of its functions despite a major structural alteration such as abnormal GCL widening.
Copyright © 2022 Karina Hernández Mercado et al.
Conflict of interest statement
The authors declare no competing financial or nonfinancial interests.
Figures




References
-
- Bouilleret V., Ridoux V., Depaulis A., Marescaux C., Nehlig A., Le Gal La Salle G. Recurrent seizures and hippocampal sclerosis following intrahippocampal kainate injection in adult mice: electroencephalography, histopathology and synaptic reorganization similar to mesial temporal lobe epilepsy. Neuroscience . 1999;89(3):717–729. doi: 10.1016/S0306-4522(98)00401-1. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
