

Plant biomechanics and resilience to environmental changes are controlled by specific lignin chemistries in each vascular cell type and morphotype
- PMID: 36215679
- PMCID: PMC9709985
- DOI: 10.1093/plcell/koac284
Plant biomechanics and resilience to environmental changes are controlled by specific lignin chemistries in each vascular cell type and morphotype
Abstract
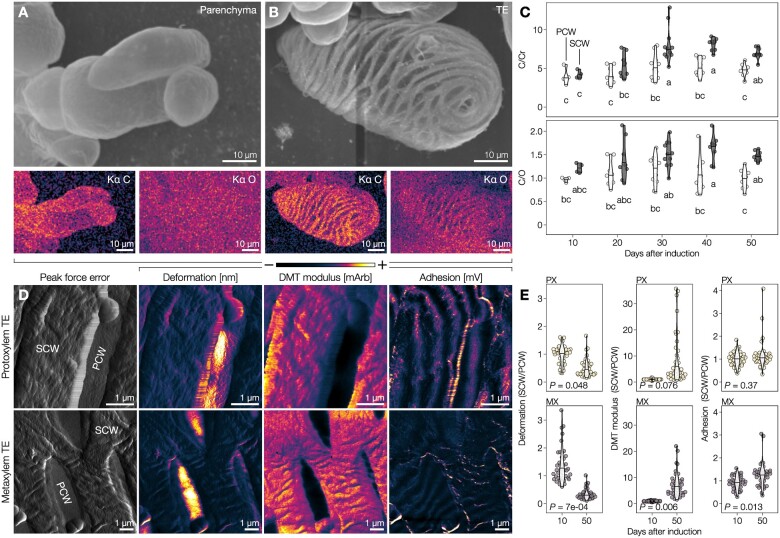
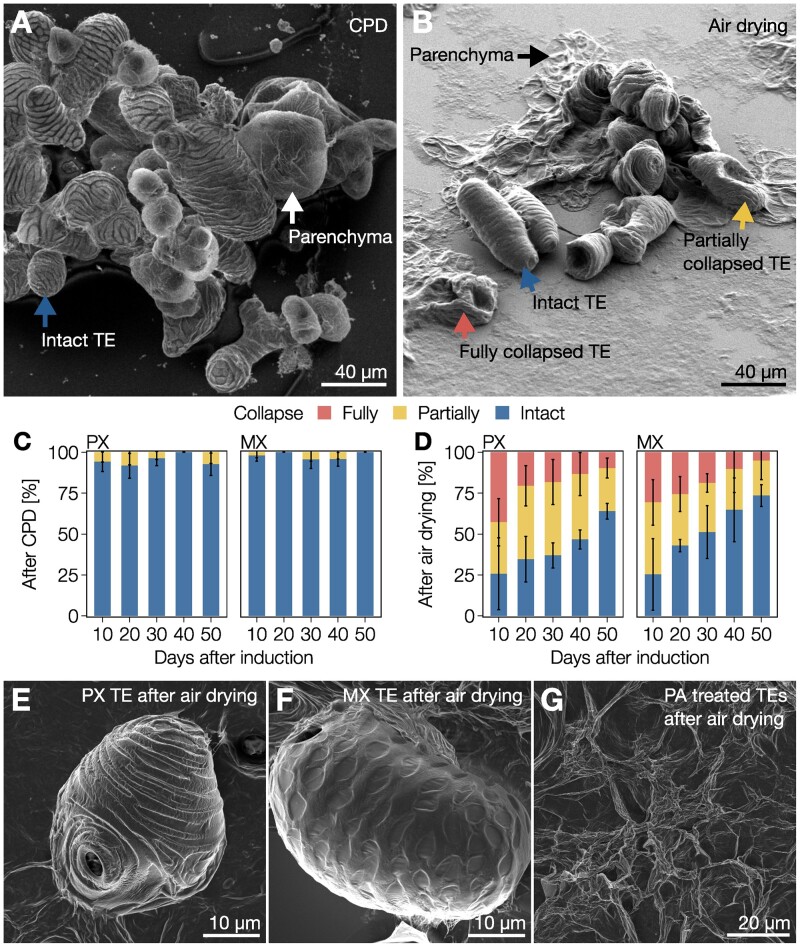
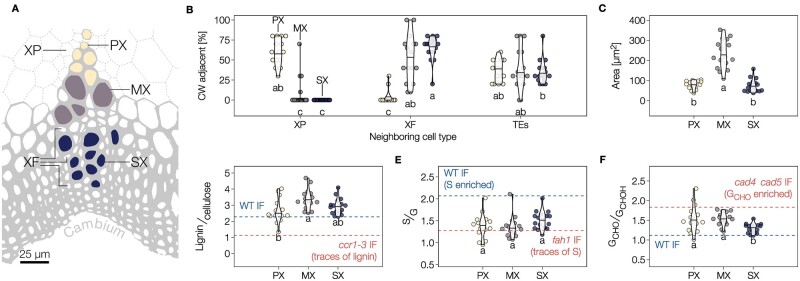
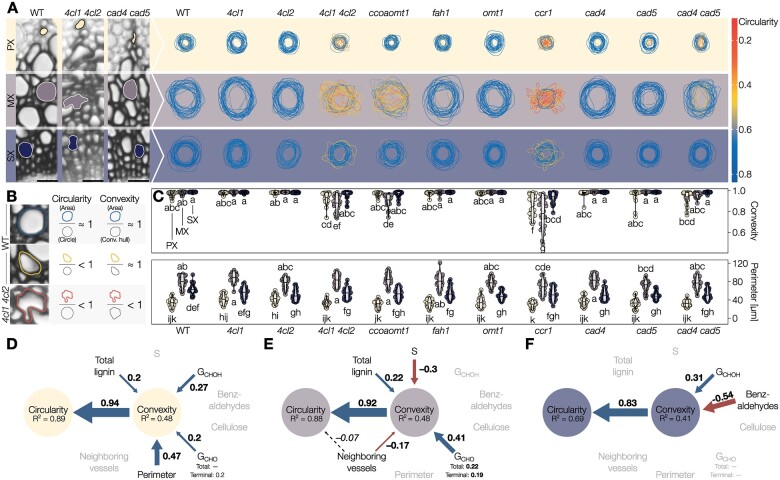
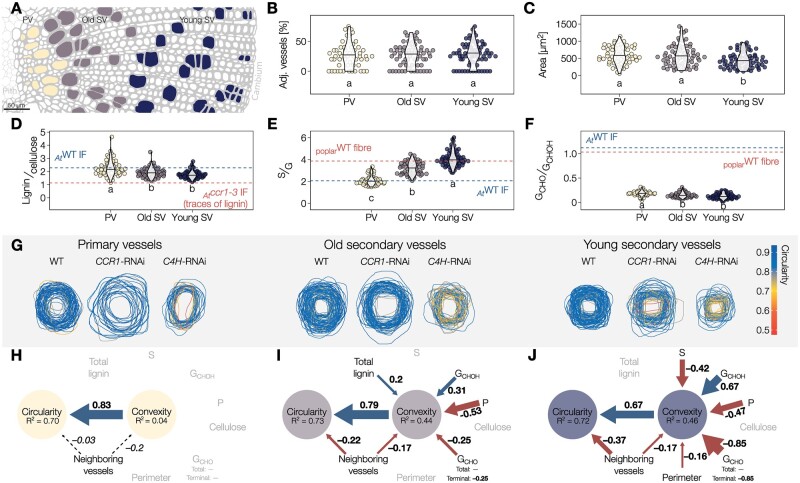
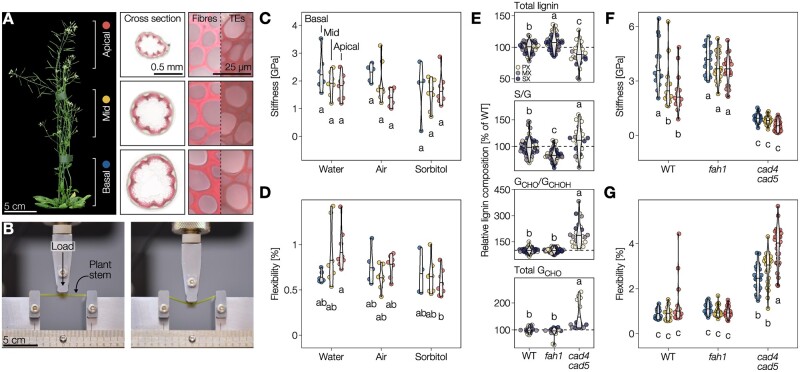
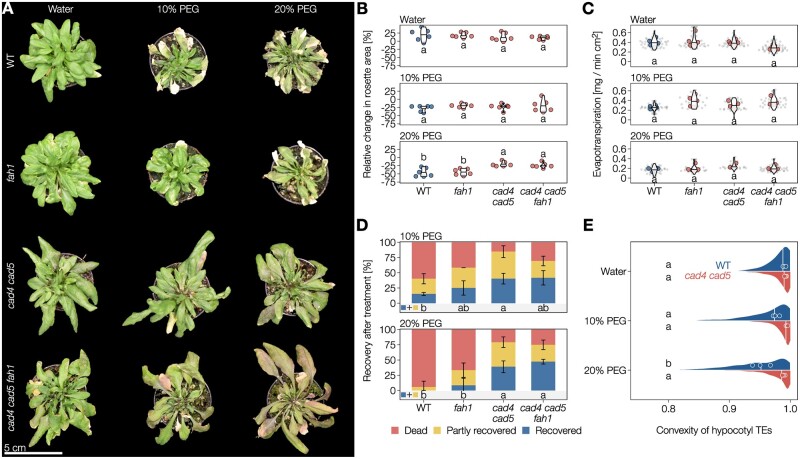
The biopolymer lignin is deposited in the cell walls of vascular cells and is essential for long-distance water conduction and structural support in plants. Different vascular cell types contain distinct and conserved lignin chemistries, each with specific aromatic and aliphatic substitutions. Yet, the biological role of this conserved and specific lignin chemistry in each cell type remains unclear. Here, we investigated the roles of this lignin biochemical specificity for cellular functions by producing single cell analyses for three cell morphotypes of tracheary elements, which all allow sap conduction but differ in their morphology. We determined that specific lignin chemistries accumulate in each cell type. Moreover, lignin accumulated dynamically, increasing in quantity and changing in composition, to alter the cell wall biomechanics during cell maturation. For similar aromatic substitutions, residues with alcohol aliphatic functions increased stiffness whereas aldehydes increased flexibility of the cell wall. Modifying this lignin biochemical specificity and the sequence of its formation impaired the cell wall biomechanics of each morphotype and consequently hindered sap conduction and drought recovery. Together, our results demonstrate that each sap-conducting vascular cell type distinctly controls their lignin biochemistry to adjust their biomechanics and hydraulic properties to face developmental and environmental constraints.
© The Author(s) 2022. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
Similar articles
-
Different combinations of laccase paralogs nonredundantly control the amount and composition of lignin in specific cell types and cell wall layers in Arabidopsis.Plant Cell. 2023 Feb 20;35(2):889-909. doi: 10.1093/plcell/koac344. Plant Cell. 2023. PMID: 36449969 Free PMC article.
-
Cooperative lignification of xylem tracheary elements.Plant Signal Behav. 2015;10(4):e1003753. doi: 10.1080/15592324.2014.1003753. Plant Signal Behav. 2015. PMID: 25761224 Free PMC article.
-
Trends in lignin modification: a comprehensive analysis of the effects of genetic manipulations/mutations on lignification and vascular integrity.Phytochemistry. 2002 Oct;61(3):221-94. doi: 10.1016/s0031-9422(02)00211-x. Phytochemistry. 2002. PMID: 12359514 Review.
-
Bulk and In Situ Quantification of Coniferaldehyde Residues in Lignin.Methods Mol Biol. 2024;2722:201-226. doi: 10.1007/978-1-0716-3477-6_14. Methods Mol Biol. 2024. PMID: 37897609
-
Transcriptional networks for lignin biosynthesis: more complex than we thought?Trends Plant Sci. 2011 Apr;16(4):227-33. doi: 10.1016/j.tplants.2010.12.005. Epub 2011 Jan 10. Trends Plant Sci. 2011. PMID: 21227733 Review.
Cited by
-
Different combinations of laccase paralogs nonredundantly control the amount and composition of lignin in specific cell types and cell wall layers in Arabidopsis.Plant Cell. 2023 Feb 20;35(2):889-909. doi: 10.1093/plcell/koac344. Plant Cell. 2023. PMID: 36449969 Free PMC article.
-
Top five unanswered questions in plant cell surface research.Cell Surf. 2024 Feb 13;11:100121. doi: 10.1016/j.tcsw.2024.100121. eCollection 2024 Jun. Cell Surf. 2024. PMID: 38405175 Free PMC article. Review.
-
Are leaf anatomical traits strong predictors of litter decomposability? Evidence from upper Andean tropical species along a forest successional gradient.Oecologia. 2025 Jun 24;207(7):110. doi: 10.1007/s00442-025-05739-8. Oecologia. 2025. PMID: 40555843 Free PMC article.
-
Analysis of the Mechanisms Underlying the Specificity of the Variation Potential Induced by Different Stimuli.Plants (Basel). 2024 Oct 16;13(20):2896. doi: 10.3390/plants13202896. Plants (Basel). 2024. PMID: 39458843 Free PMC article.
-
Modulation of lignin biosynthesis for drought tolerance in plants.Front Plant Sci. 2023 Apr 20;14:1116426. doi: 10.3389/fpls.2023.1116426. eCollection 2023. Front Plant Sci. 2023. PMID: 37152118 Free PMC article. Review.
References
-
- Abraham MJ, Murtola T, Schulz R, Páll S, Smith JC, Hess B, Lindahl E (2015) GROMACS: high performance molecular simulations through multi-level parallelism from laptops to supercomputers. SoftwareX 1–2: 19–25
-
- Amrhein N, Frank G, Lemm G, Luhmann HB (1983) Inhibition of lignin formation by L-alpha-aminooxy-beta-phenylpropionic acid, an inhibitor of phenylalanine ammonia-lyase. Eur J Cell Biol 29: 139–144 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
