

Sequential action of a tRNA base editor in conversion of cytidine to pseudouridine
- PMID: 36220828
- PMCID: PMC9553926
- DOI: 10.1038/s41467-022-33714-x
Sequential action of a tRNA base editor in conversion of cytidine to pseudouridine
Abstract
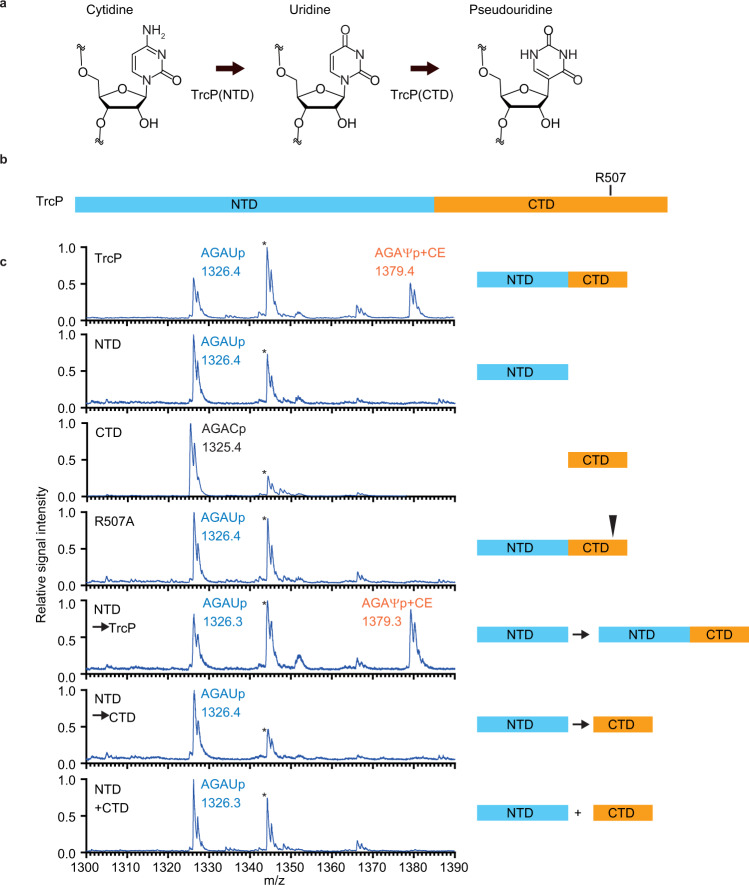
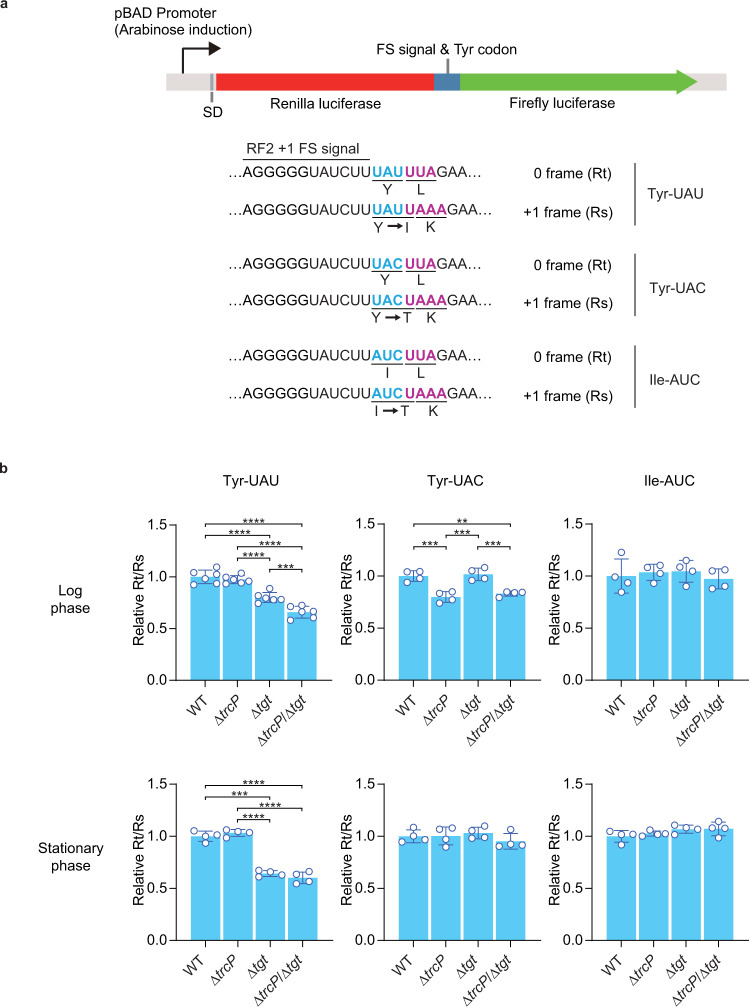
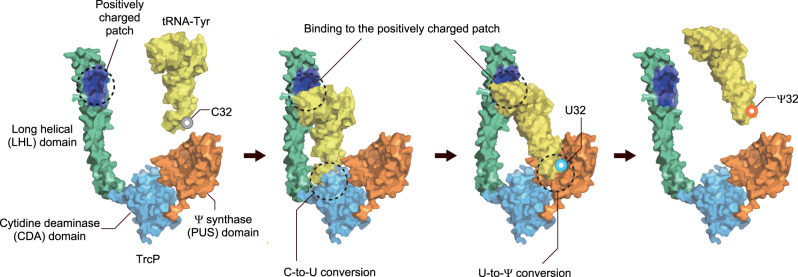
Post-transcriptional RNA editing modulates gene expression in a condition-dependent fashion. We recently discovered C-to-Ψ editing in Vibrio cholerae tRNA. Here, we characterize the biogenesis, regulation, and functions of this previously undescribed RNA editing process. We show that an enzyme, TrcP, mediates the editing of C-to-U followed by the conversion of U to Ψ, consecutively. AlphaFold-2 predicts that TrcP consists of two globular domains (cytidine deaminase and pseudouridylase) and a long helical domain. The latter domain tethers tRNA substrates during both the C-to-U editing and pseudouridylation, likely enabling a substrate channeling mechanism for efficient catalysis all the way to the terminal product. C-to-Ψ editing both requires and suppresses other modifications, creating an interdependent network of modifications in the tRNA anticodon loop that facilitates coupling of tRNA modification states to iron availability. Our findings provide mechanistic insights into an RNA editing process that likely promotes environmental adaptation.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures




Similar articles
-
ADATs: roles in tRNA editing and relevance to disease.Acta Biochim Biophys Sin (Shanghai). 2024 Jul 22;57(1):73-83. doi: 10.3724/abbs.2024125. Acta Biochim Biophys Sin (Shanghai). 2024. PMID: 39034823 Free PMC article. Review.
-
Role of editing in plant mitochondrial transfer RNAs.Gene. 2002 Mar 6;286(1):21-4. doi: 10.1016/s0378-1119(01)00817-4. Gene. 2002. PMID: 11943456
-
Major identity determinants for enzymatic formation of ribothymidine and pseudouridine in the T psi-loop of yeast tRNAs.J Mol Biol. 1997 Dec 12;274(4):505-18. doi: 10.1006/jmbi.1997.1417. J Mol Biol. 1997. PMID: 9417931
-
An adenosine-to-inosine tRNA-editing enzyme that can perform C-to-U deamination of DNA.Proc Natl Acad Sci U S A. 2007 May 8;104(19):7821-6. doi: 10.1073/pnas.0702394104. Epub 2007 May 1. Proc Natl Acad Sci U S A. 2007. PMID: 17483465 Free PMC article.
-
Enzymatic conversion of cytidine to lysidine in anticodon of bacterial isoleucyl-tRNA--an alternative way of RNA editing.Trends Biochem Sci. 2004 Apr;29(4):165-8. doi: 10.1016/j.tibs.2004.02.009. Trends Biochem Sci. 2004. PMID: 15124629 Review. No abstract available.
Cited by
-
A tRNA modification in Mycobacterium tuberculosis facilitates optimal intracellular growth.bioRxiv [Preprint]. 2023 Jun 9:2023.02.20.529267. doi: 10.1101/2023.02.20.529267. bioRxiv. 2023. Update in: Elife. 2023 Sep 27;12:RP87146. doi: 10.7554/eLife.87146. PMID: 36865327 Free PMC article. Updated. Preprint.
-
Aminoglycoside tolerance in Vibrio cholerae engages translational reprogramming associated with queuosine tRNA modification.Elife. 2025 Jan 6;13:RP96317. doi: 10.7554/eLife.96317. Elife. 2025. PMID: 39761105 Free PMC article.
-
RudS: bacterial desulfidase responsible for tRNA 4-thiouridine de-modification.Nucleic Acids Res. 2024 Sep 23;52(17):10543-10562. doi: 10.1093/nar/gkae716. Nucleic Acids Res. 2024. PMID: 39166491 Free PMC article.
-
Molecular Coping Mechanisms: Reprogramming tRNAs To Regulate Codon-Biased Translation of Stress Response Proteins.Acc Chem Res. 2023 Dec 5;56(23):3504-3514. doi: 10.1021/acs.accounts.3c00572. Epub 2023 Nov 22. Acc Chem Res. 2023. PMID: 37992267 Free PMC article.
-
ADATs: roles in tRNA editing and relevance to disease.Acta Biochim Biophys Sin (Shanghai). 2024 Jul 22;57(1):73-83. doi: 10.3724/abbs.2024125. Acta Biochim Biophys Sin (Shanghai). 2024. PMID: 39034823 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
