

doi: 10.1093/plcell/koac303.
Climate change challenges, plant science solutions
Affiliations
- PMID: 36222573
- PMCID: PMC9806663
- DOI: 10.1093/plcell/koac303
Item in Clipboard
Climate change challenges, plant science solutions
Plant Cell.
.
Abstract
Climate change is a defining challenge of the 21st century, and this decade is a critical time for action to mitigate the worst effects on human populations and ecosystems. Plant science can play an important role in developing crops with enhanced resilience to harsh conditions (e.g. heat, drought, salt stress, flooding, disease outbreaks) and engineering efficient carbon-capturing and carbon-sequestering plants. Here, we present examples of research being conducted in these areas and discuss challenges and open questions as a call to action for the plant science community.
© The Author(s) 2022. Published by Oxford University Press on behalf of American Society of Plant Biologists.
Figures
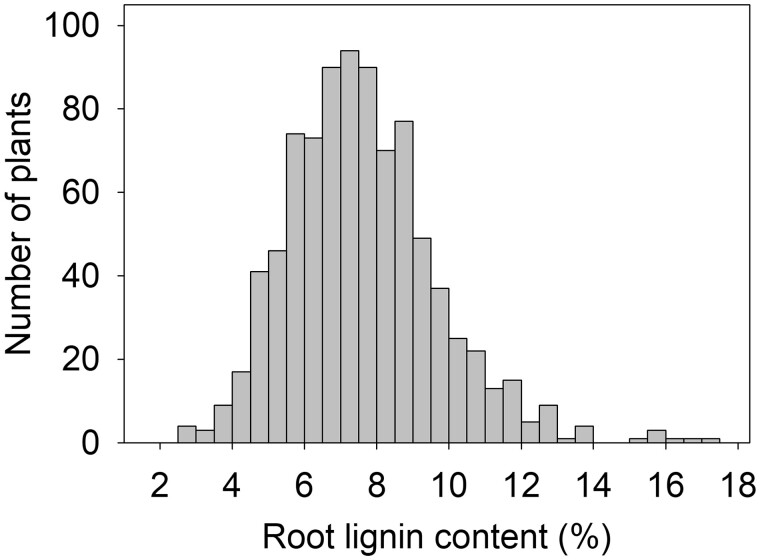
Quantitative variation in lignin content in maize root systems from a field study of 358 maize inbred lines. A description of the experiment can be found in Woods et al. (2022).
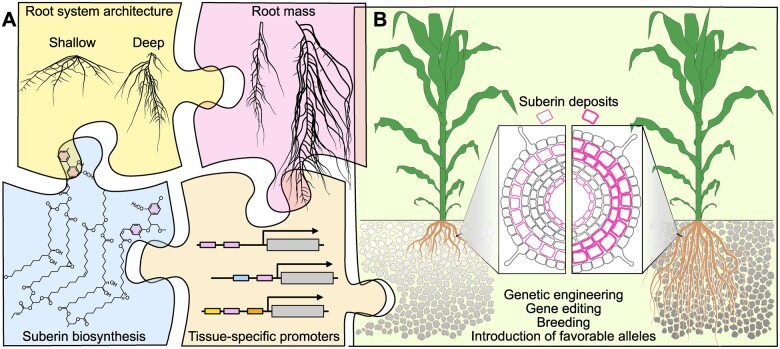
Toward an ideal carbon-capturing crop plant. A, The ideal plant should accumulate suberin in the cell wall of its root cells and form a vast and deep root system. To realize this goal, the existing literature and experimental evidence are curated to look for candidate genes affecting root system architecture and root mass. This information is combined with root-specific promoters and suberin biosynthetic genes. B, The ideal plant is created by capitalizing on both classical (breeding) and more recent (genome editing, genetic engineering) approaches to introduce favorable alleles and genes that will increase root biomass and transgenes that will increase the deposition of suberin in the root. In addition to trapping more carbon, these ideal plants will replenish carbon-depleted soils with degradation-recalcitrant carbon polymers (indicated by the darker color of the soil on the right). Figure credit: P. Salomé.

Examples of wide hybridization and direct domestication to develop perennial grains. The wild perennial Sorghum halepense (A) was hybridized with the domestic species Sorghum bicolor (B) and selective breeding of the progeny produced lines with intermediate head and seed size (C) and the ability to regrow from underground rhizomes (D). In an example of direct domestication, the mostly wild grass Thinopyrum intermedium can be harvested with conventional equipment (E) and cleaned to obtain a human-edible grain (F) that has properties similar to wheat, as seen in this loaf made with an 80/20 blend of wheat and Th. intermedium flour (G). Domesticated Th. intermedium types now possess domestication traits, such as shatter resistance (H, at right).
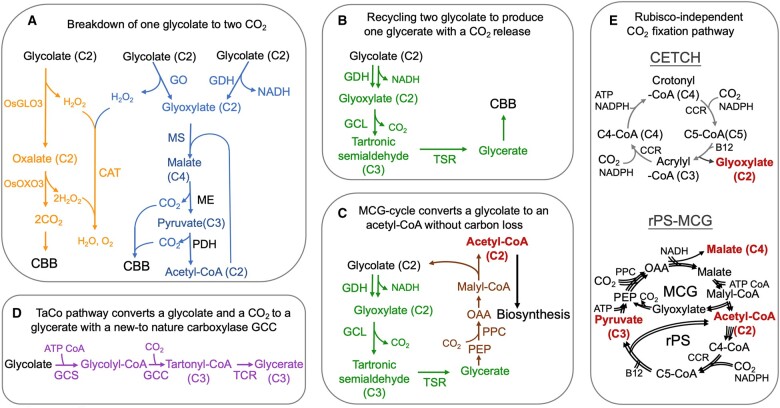
Synthetic biology approaches for recycling photorespiration and CO2-fixation pathways. A and B, Photorespiration engineered for breakdown of one glycolate to two CO2 molecules (A) or conversion of two glycolate to one glycerate plus one CO2 molecule (B). The CO2 released in the chloroplast is recycled back to the CBB cycle for carbon reassimilation (Maier et al., 2012; Shen et al., 2019; South et al., 2019). C, The MCG-cycle engineered to convert glycolate to acetyl-CoA without carbon loss (Yu et al., 2018b). D, Creation of a new enzyme, such as glycolyl-CoA carboxylase, to achieve glycolate recycling to produce glycerate with input of ATP and an additional CO2 molecule (Scheffen et al., 2021). E, Rubisco-independent CETCH and rPS-MCG synthetic CO2-fixation pathways (Luo et al., 2022).
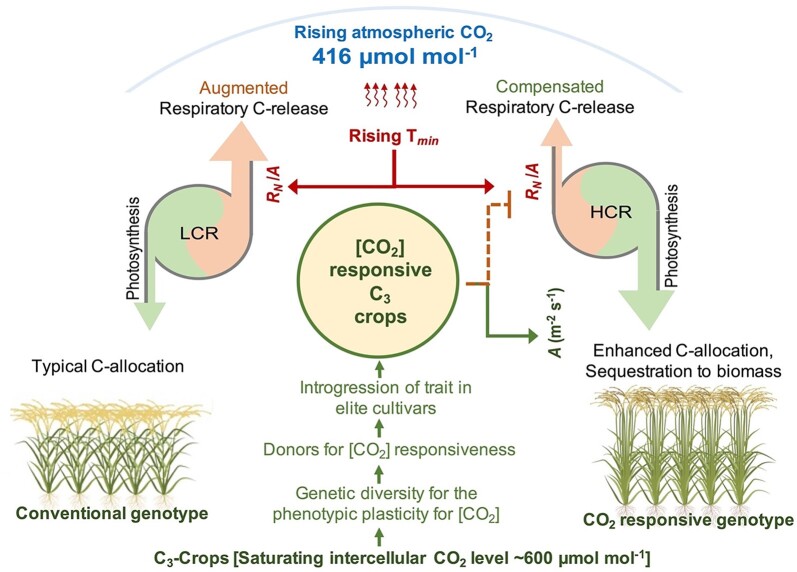
Schematic diagram showing average annual atmospheric [CO2] level for 2021 and the effect of rising night temperature (Tmin) on rice productivity by enhanced respiration: photosynthesis ratio (RN/A) resulting in augmented release of carbon at the cost of biomass and yield in conventional genotypes. On the contrary, introgression of CO2-responsiveness trait in C3 crops facilitates enhanced carbon sequestration and allocation of additional carbon into biomass, and compensating Tmin-induced carbon losses. LCR, least CO2-responsive; HCR, high CO2-responsive.
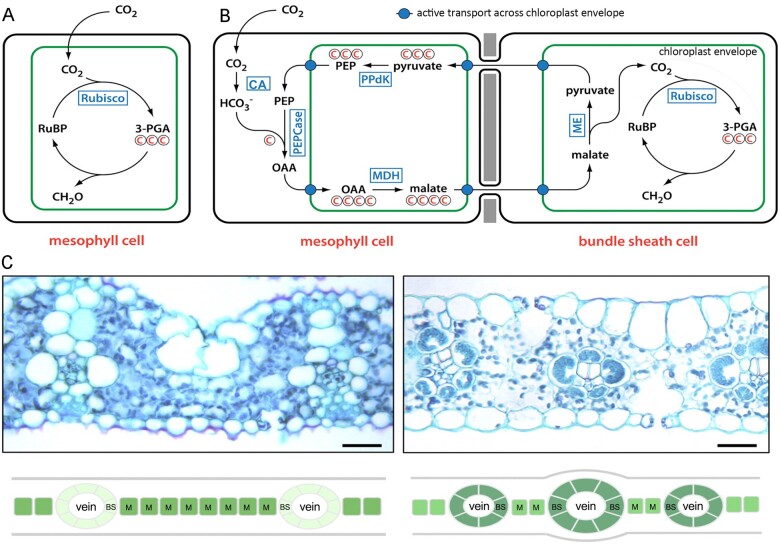
Schematics of C3 CBB and NADP-ME C4 Cycles. A, CBB C3 cycle. B, NADP-ME C4 cycle. C, Transverse leaf sections and corresponding schematics of C3 rice (left) and C4 maize (right). Bars = 30 μm. Adapted from Langdale (2011), Figures 1 and 3.
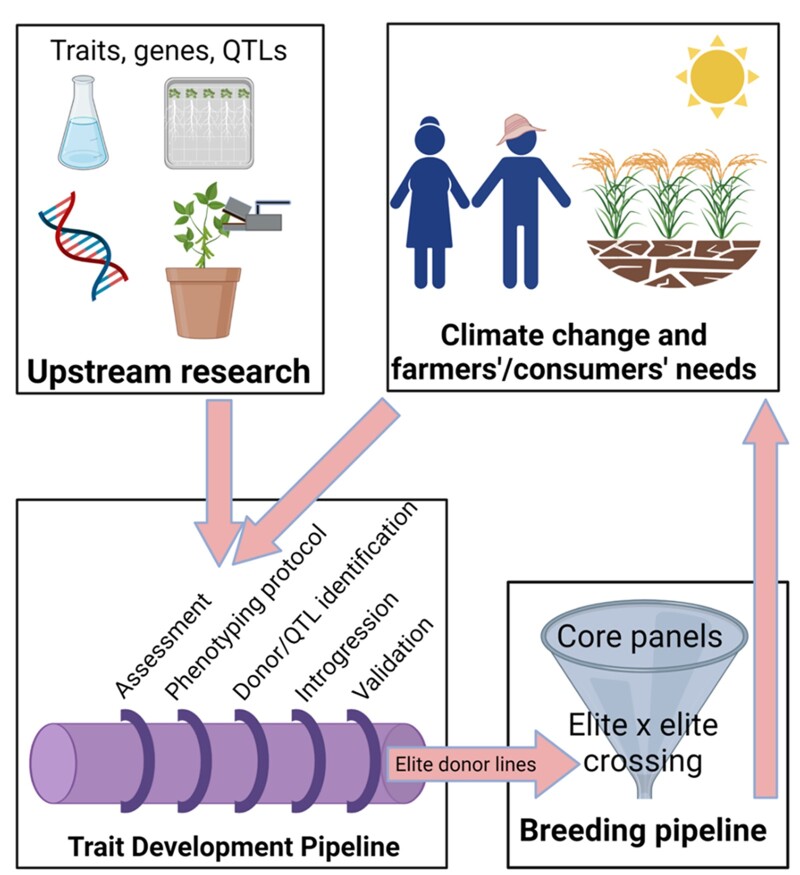
The Trait Development Pipeline for delivery of valuable stress tolerance traits/genes/QTLs from upstream research into the elite breeding pool for improvement of crop productivity under climate change. The Trait Development Pipeline is organized into six stages: 1) guidelines for prioritizing traits (assessment), 2) defining standards for phenotyping protocols, 3) identifying donors and QTL (including refining marker quality metrics), 4) introgressing and 5) validating traits/genes/QTLs into elite genetic backgrounds to develop the elite donor lines that are 6) handed to the breeding program for crossing. Those elite donor lines will then be systematically crossed and tested in target environments where climate change is increasingly affecting the degree of abiotic stress affecting crop production. Created with BioRender.com
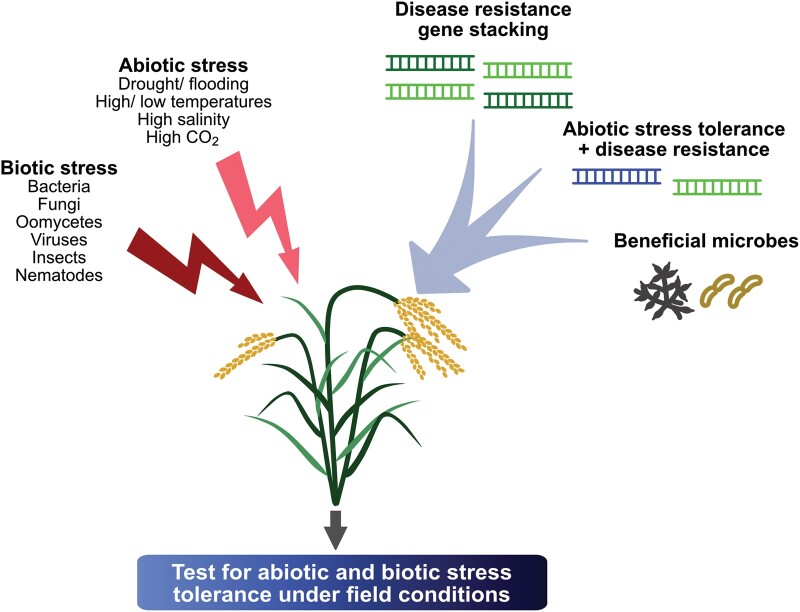
Development of crops with enhanced resilience to abiotic and biotic stress. Crops are exposed to a variety of stresses. Abiotic stresses will intensify as the following climate conditions change: water availability, precipitation, temperatures, and atmospheric CO2 levels. Biotic stressors that plants encounter will vary, but may consist of: bacteria, fungi, oomycetes, nematodes, viruses, and insect pests. As climate change alters environmental conditions and plant-pathogen interactions, strategies to develop more climate-ready and disease-resistant crop varieties include breeding or genome engineering approaches with stacking disease resistance genes, stacking climate tolerance and disease resistance genes, and/or addition of beneficial microbes (see text for examples).
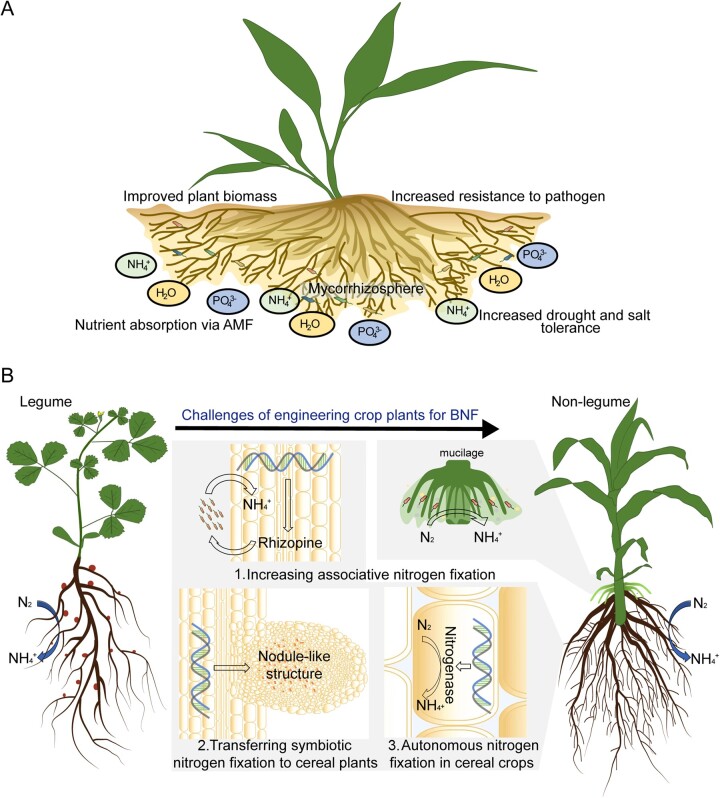
Mycorrhizal symbiosis and N self-fertilizing crops. A, Positive effects of mycorrhizal symbiosis. The mycorrhizal hyphal network forms a mycorrhizosphere (light brown) which can enlarge the plant nutrient absorption area and supply a convenient zone for root-related microbes. Benefits from mycorrhizal symbiosis include increased tolerance or resistance to abiotic or biotic stresses. B, Three steps to develop N self-fertilizing cereal crops to enhance climate change resilience. (1) Increasing associative N fixation. The mucilage (light green) is rich in carbohydrates and harbors abundant diazotrophic microbiota (pink). Engineered cereal plants (such as maize) have the ability to produce rhizophine, which can be perceived by engineered diazotrophs (orange). (2) Transferring symbiotic N fixation to cereal plants. Cereal crops are engineered for symbiotic N fixation by expressing the chimeric receptors perceiving rhizobia signals and overexpressing key symbiotic regulators (CSSP genes, CRE1, etc.) and nodule development genes (SCR-SHR, LBD16, etc.) to form nodule-like structures. (3) Autonomous N fixation in cereal crops. The ideal plant which could assimilate N2 into ammonium is created by overexpressing rhizobial N fixation genes in plant cells.
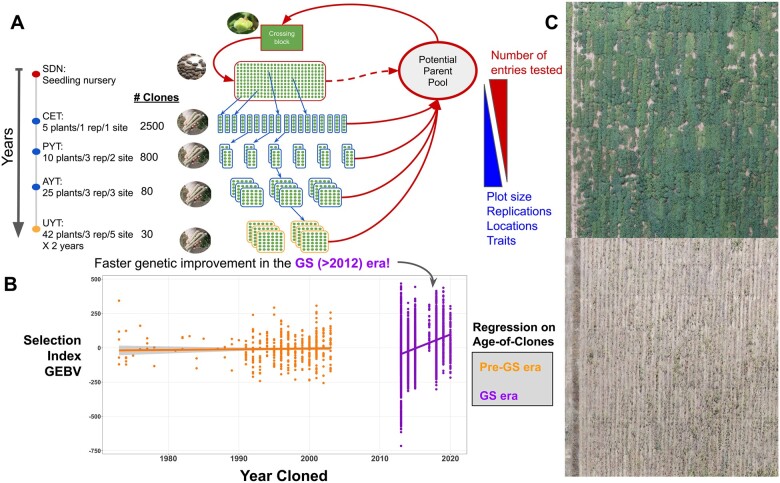
Genomic selection (GS) in a cassava breeding program. A, Each breeding cycle begins with a crossing block trial where seeds are generated. The first evaluation, a seedling nursery (SDN) usually involves >10K plants, but cassava does not produce storage roots when planted from seed and no yield data is collected. After 12 months, seedlings are cloned (5–10 cuttings/plant) into their first single-row, unreplicated clonal evaluation trial (CET) followed by at least three stages of yield trials (preliminary [PYT], advanced [AYT], and uniform [UYT]). All lines entering CET are genotyped genome-wide; sometimes this is done during the seedling nursery. As a result, genomic prediction enables selection of new parents for crossing even as early as the SDN (dashed red arrow). B, GS has resulted in demonstrable acceleration in the rate of genetic improvement since initiation in 2012. Results shown are from the IITA GS population. The genomically predicted performance of GS-era (purple) and historical (yellow) clones relative to a multi-trait selection index (y-axis) is plotted against the year when each clone was first generated (x-axis). C, Field trial showing variability for one of the major future challenges to cassava: drought. The top image shows plants 3 months after planting, under irrigation at Petrolina (Pernambuco, Brazil). The bottom image shows plants 3 months later under water deficit.
References
-
- Agbona A, Teare B, Ruiz-Guzman H, Dobreva ID, Everett ME, Adams T, Montesinos-Lopez OA, Kulakow PA, Hays DB (2021) Prediction of root biomass in cassava based on ground penetrating radar phenomics. Remote Sens 13: 4908
-
- Aggarwal P, Vyas S, Thornton P, Campbell BM (2019) How much does climate change add to the challenge of feeding the planet this century? Environ Res Lett 14: 043001
-
- Ahmadi J, Pour-Aboughadareh A, Fabriki-Ourang S, Mehrabi AA, Siddique KH (2018) Screening wild progenitors of wheat for salinity stress at early stages of plant growth: insight into potential sources of variability for salinity adaptation in wheat. Crop Pasture Sci 69: 649–658
-
- Ainsworth EA, Long SP (2021) 30 years of free-air carbon dioxide enrichment (FACE): What have we learned about future crop productivity and its potential for adaptation? Global Change Biol 27: 27–49 - PubMed
-
- Albritton DL, Meira Filho LG, Cubasch U, Dai X, Ding Y, Griggs DJ, Hewitson B, Houghton JT, Isaksen I, Karl T. et al. (2001) Technical Summary. In Houghton JT, Ding Y, Griggs DJ, Noguer M, van der Linden PJ, Dai X, Maskell K, Johnson CA, eds, Climate Change 2001: The scientific basis. Contribution of working group I to the third assessment report of the intergovernmental panel on climate change, Cambridge University Press, Cambridge, United Kingdom and New York, NY, USA, pp. 881
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
