

Synaptic ensembles between raphe and D1R-containing accumbens shell neurons underlie postisolation sociability in males
- PMID: 36223467
- PMCID: PMC9555785
- DOI: 10.1126/sciadv.abo7527
Synaptic ensembles between raphe and D1R-containing accumbens shell neurons underlie postisolation sociability in males
Abstract
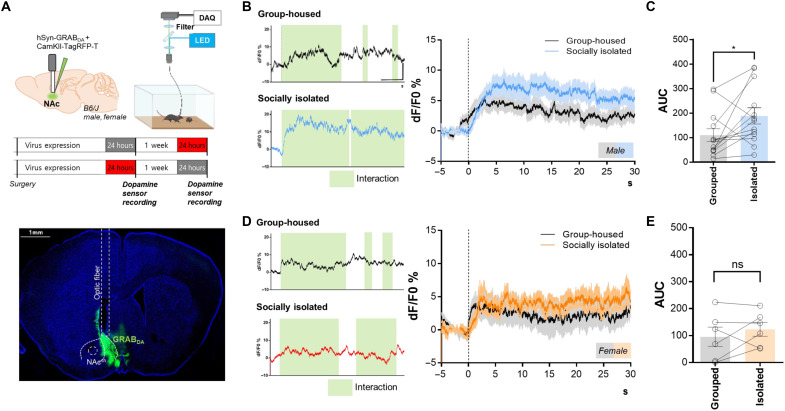
Social animals expend considerable energy to maintain social bonds throughout their life. Male and female mice show sexually dimorphic behaviors, yet the underlying neural mechanisms of sociability and their dysregulation during social disconnection remain unknown. Dopaminergic neurons in dorsal raphe nucleus (DRNTH) is known to contribute to a loneliness-like state and modulate sociability. We identified that activated subpopulations in DRNTH and nucleus accumbens shell (NAcsh) during 24 hours of social isolation underlie the increase in isolation-induced sociability in male but not in female mice. This effect was reversed by chemogenetically and optogenetically inhibiting the DRNTH-NAcsh circuit. Moreover, synaptic connectivity among the activated neuronal ensembles in this circuit was increased, primarily in D1 receptor-expressing neurons in NAcsh. The increase in synaptic density functionally correlated with elevated dopamine release into NAcsh. Overall, specific synaptic ensembles in DRNTH-NAcsh mediate sex differences in isolation-induced sociability, indicating that sex-dependent circuit dynamics underlie the expression of sexually dimorphic behaviors.
Figures





Similar articles
-
Dopamine D1 receptor in the NAc shell is involved in delayed emergence from isoflurane anesthesia in aged mice.Brain Behav. 2021 Jan;11(1):e01913. doi: 10.1002/brb3.1913. Epub 2020 Oct 22. Brain Behav. 2021. PMID: 33094567 Free PMC article.
-
Differential roles of dopamine D1 and D2 receptor-containing neurons of the nucleus accumbens shell in behavioral sensitization.J Neurochem. 2015 Dec;135(6):1232-41. doi: 10.1111/jnc.13380. Epub 2015 Oct 28. J Neurochem. 2015. PMID: 26442961
-
Changes in Appetitive Associative Strength Modulates Nucleus Accumbens, But Not Orbitofrontal Cortex Neuronal Ensemble Excitability.J Neurosci. 2017 Mar 22;37(12):3160-3170. doi: 10.1523/JNEUROSCI.3766-16.2017. Epub 2017 Feb 17. J Neurosci. 2017. PMID: 28213443 Free PMC article.
-
Negative consequences of early-life adversity on substance use as mediated by corticotropin-releasing factor modulation of serotonin activity.Neurobiol Stress. 2018 Aug 7;9:29-39. doi: 10.1016/j.ynstr.2018.08.001. eCollection 2018 Nov. Neurobiol Stress. 2018. PMID: 30151419 Free PMC article. Review.
-
Reward processing by the dorsal raphe nucleus: 5-HT and beyond.Learn Mem. 2015 Aug 18;22(9):452-60. doi: 10.1101/lm.037317.114. Print 2015 Sep. Learn Mem. 2015. PMID: 26286655 Free PMC article. Review.
Cited by
-
The nucleus accumbens in reward and aversion processing: insights and implications.Front Behav Neurosci. 2024 Aug 9;18:1420028. doi: 10.3389/fnbeh.2024.1420028. eCollection 2024. Front Behav Neurosci. 2024. PMID: 39184934 Free PMC article. Review.
-
Differential brain activation and network connectivity in social interactions presence and absence of physical contact.Commun Biol. 2025 Jul 2;8(1):986. doi: 10.1038/s42003-025-08417-w. Commun Biol. 2025. PMID: 40603622 Free PMC article.
-
Modulation of social valence by insular cortex activity during acute social isolation in mice.Mol Brain. 2025 Jul 15;18(1):62. doi: 10.1186/s13041-025-01236-4. Mol Brain. 2025. PMID: 40665452 Free PMC article.
-
Separable dorsal raphe dopamine projections mimic the facets of a loneliness-like state.Elife. 2025 Aug 1;14:RP105955. doi: 10.7554/eLife.105955. Elife. 2025. PMID: 40747785 Free PMC article.
-
Increased social interaction in Shank2-deficient mice following acute social isolation.Mol Brain. 2023 Apr 15;16(1):35. doi: 10.1186/s13041-023-01025-x. Mol Brain. 2023. PMID: 37061705 Free PMC article.
References
-
- Baumeister R. F., Leary M. R., The need to belong: Desire for interpersonal attachments as a fundamental human motivation. Psychol. Bull. 117, 497–529 (1995). - PubMed
-
- Van Loo P. L. P., de Groot A. C., Van Zutphen B. F. M., Baumans V., Do male mice prefer or avoid each other’s company? influence of hierarchy, kinship, and familiarity. J. Appl. Anim. Welf. Sci. 4, 91–103 (2001).
-
- Thoenig R. H., Solitary confinement-punishment within the letter of the law, or psychological torture. Wisconsin Law Rev. , 223–311 (1972).
-
- Cacioppo J. T., Hughes M. E., Waite L. J., Hawkley L. C., Thisted R. A., Loneliness as a specific risk factor for depressive symptoms: Cross-sectional and longitudinal analyses. Psychol. Aging 21, 140–151 (2006). - PubMed
-
- Fone K. C. F., Porkess M. V., Behavioural and neurochemical effects of post-weaning social isolation in rodents—Relevance to developmental neuropsychiatric disorders. Neurosci. Biobehav. Rev. 32, 1087–1102 (2008). - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
