

Maternal gut microbiota mediate intergenerational effects of high-fat diet on descendant social behavior
- PMID: 36223744
- PMCID: PMC9597666
- DOI: 10.1016/j.celrep.2022.111461
Maternal gut microbiota mediate intergenerational effects of high-fat diet on descendant social behavior
Erratum in
-
Maternal gut microbiota mediate intergenerational effects of high-fat diet on descendant social behavior.Cell Rep. 2023 May 30;42(5):112498. doi: 10.1016/j.celrep.2023.112498. Epub 2023 May 2. Cell Rep. 2023. PMID: 37133995 Free PMC article. No abstract available.
Abstract
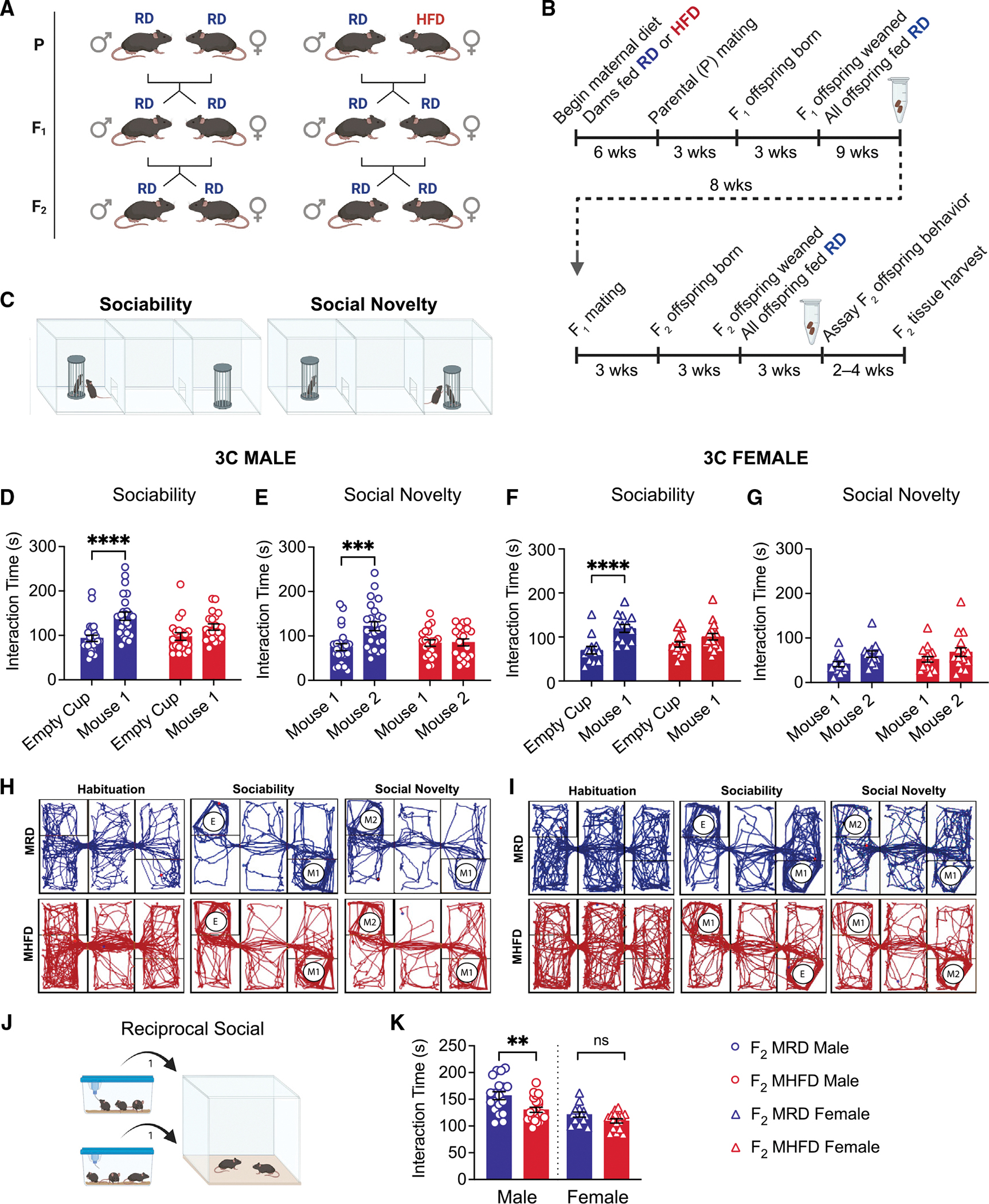
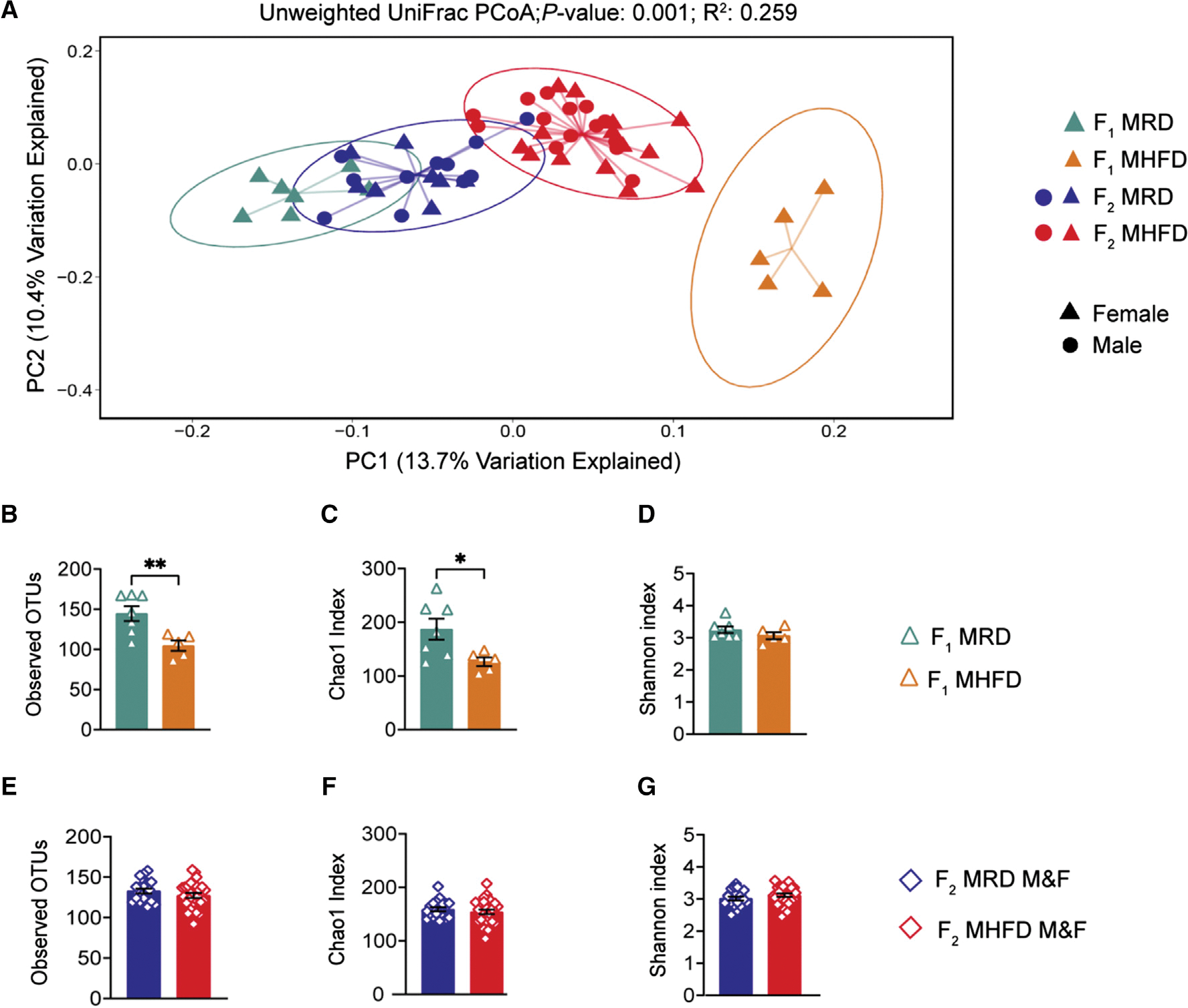
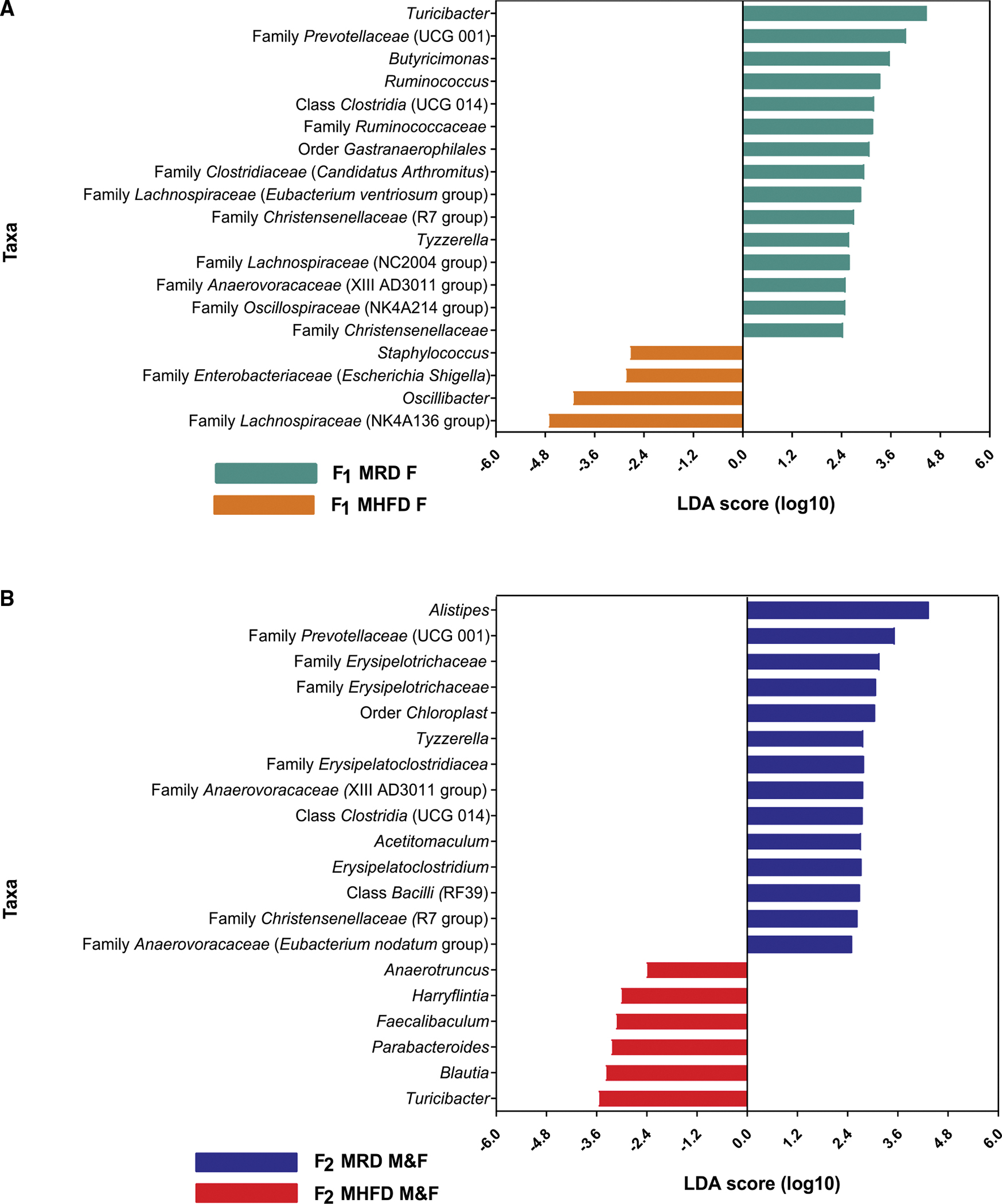
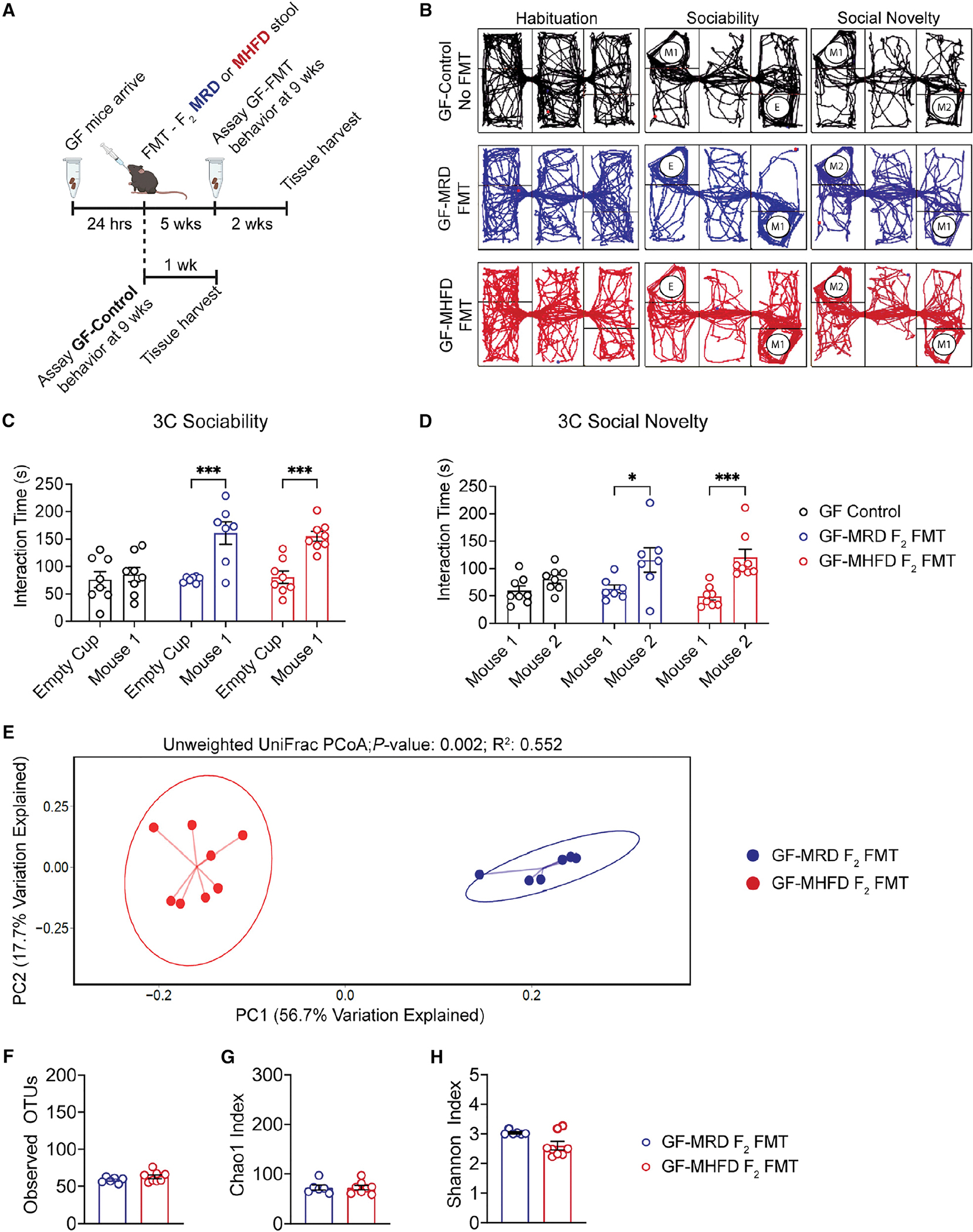
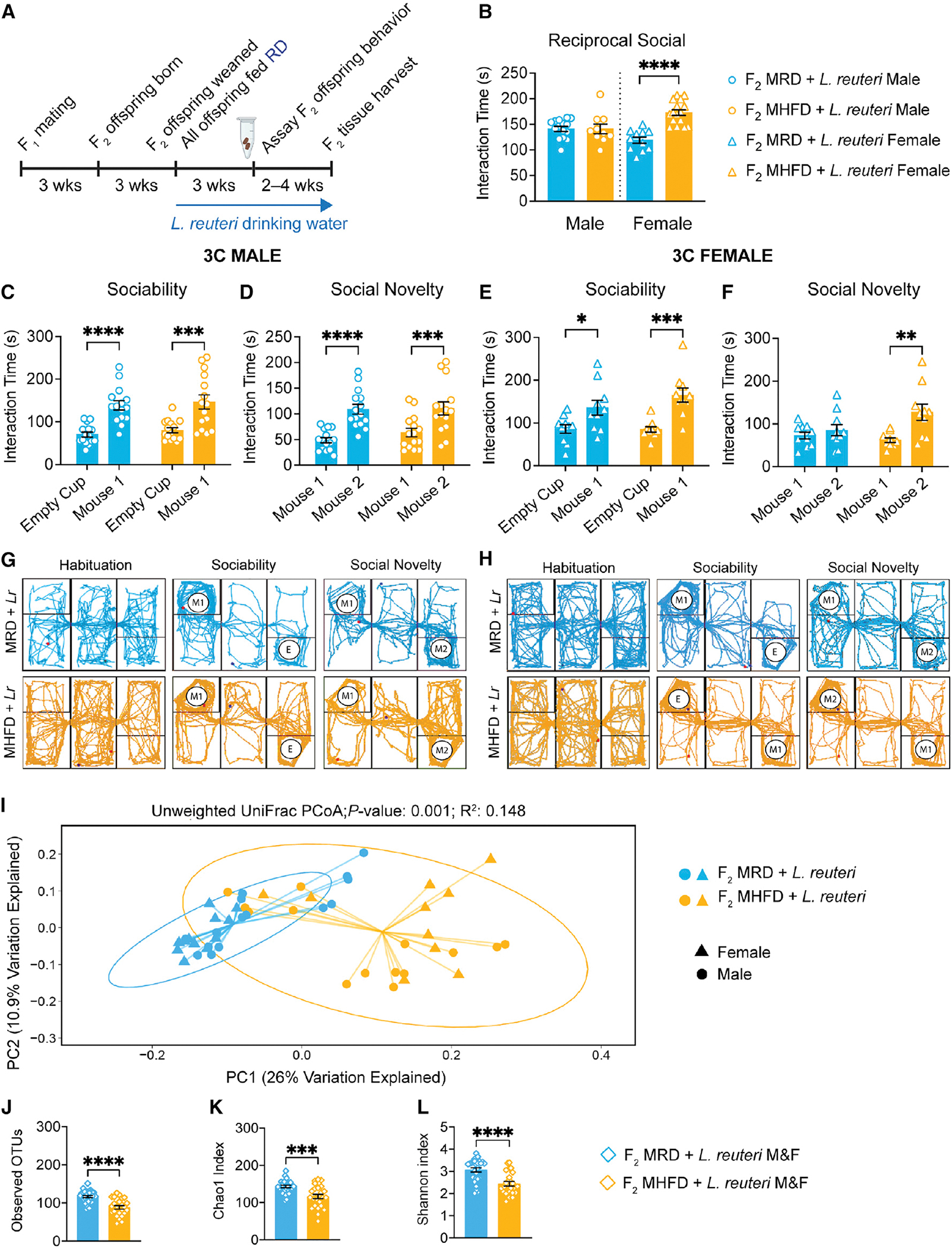
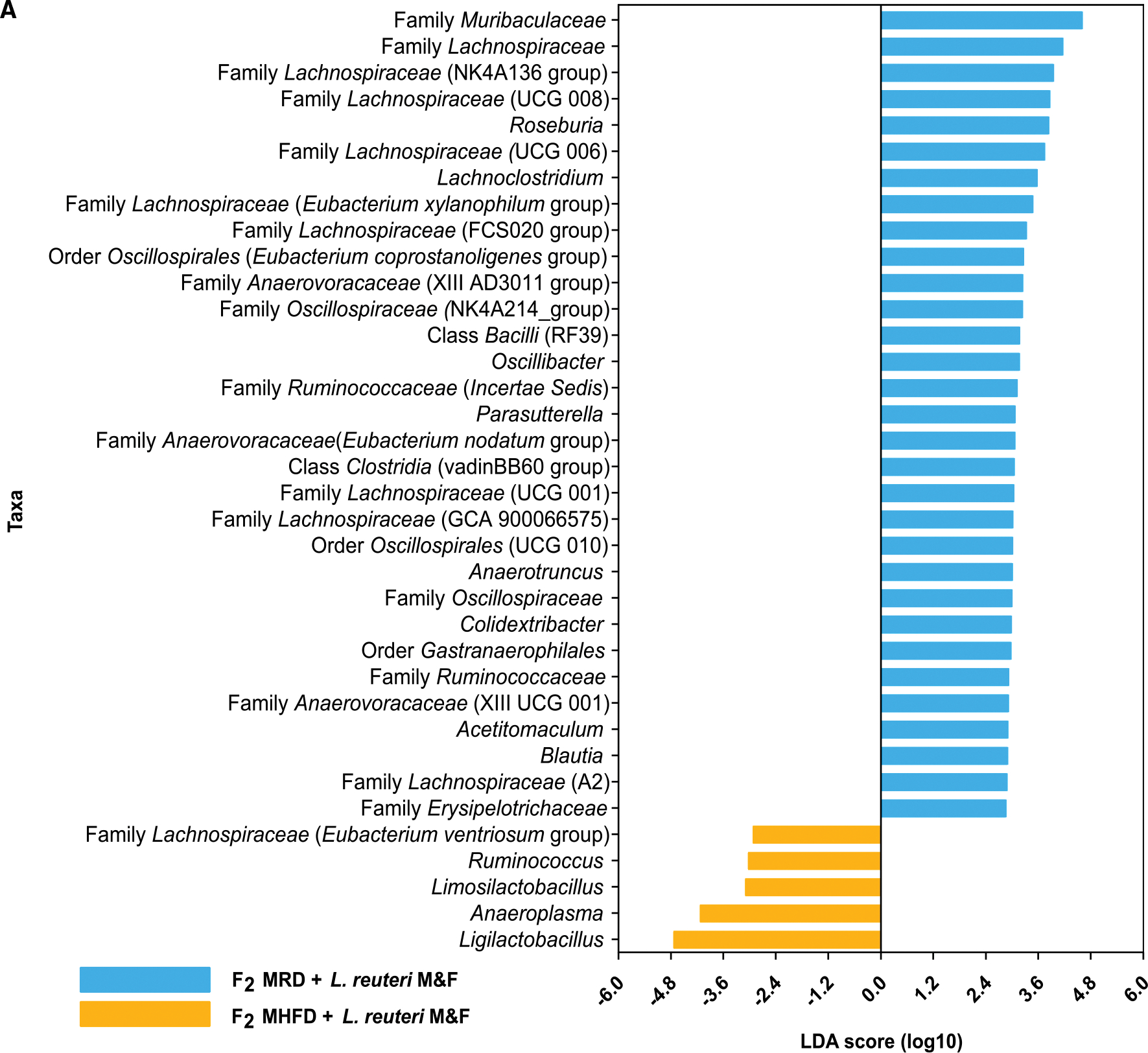
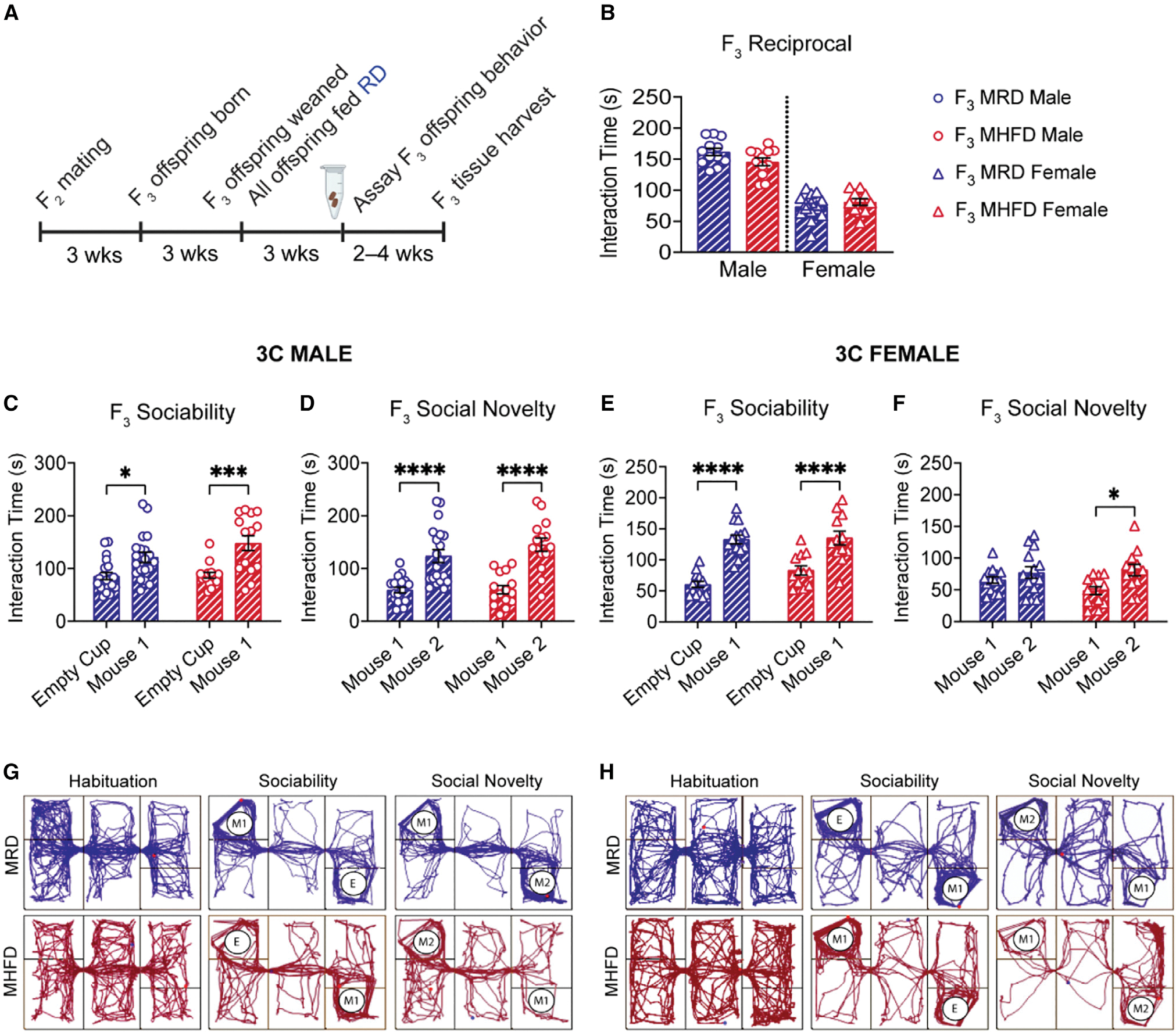
Dysbiosis of the maternal gut microbiome during pregnancy is associated with adverse neurodevelopmental outcomes. We previously showed that maternal high-fat diet (MHFD) in mice induces gut dysbiosis, social dysfunction, and underlying synaptic plasticity deficits in male offspring (F1). Here, we reason that, if HFD-mediated changes in maternal gut microbiota drive offspring social deficits, then MHFD-induced dysbiosis in F1 female MHFD offspring would likewise impair F2 social behavior. Metataxonomic sequencing reveals reduced microbial richness among female F1 MHFD offspring. Despite recovery of microbial richness among MHFD-descendant F2 mice, they display social dysfunction. Post-weaning Limosilactobacillus reuteri treatment increases the abundance of short-chain fatty acid-producing taxa and rescues MHFD-descendant F2 social deficits. L. reuteri exerts a sexually dimorphic impact on gut microbiota configuration, increasing discriminant taxa between female cohorts. Collectively, these results show multigenerational impacts of HFD-induced dysbiosis in the maternal lineage and highlight the potential of maternal microbiome-targeted interventions for neurodevelopmental disorders.
Keywords: CP: Microbiology; CP: Neuroscience; DOHaD; Limosilactobacillus reuteri; intergenerational; maternal diet; microbiome; neurodevelopment; probiotics; social behavior.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests S.A.B. is an inventor on a patent granted to Baylor College of Medicine related to the use of Limosilactobacillus reuteri for treating disorders characterized by social dysfunction (US Patent No. 11135252). The authors declare no other competing interests.
Figures
References
-
- Baron-Cohen S, Knickmeyer RC, and Belmonte MK (2005). Sex differences in the brain: implications for explaining autism. Science 310, 819–823. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
