

A generalised approach to the study and understanding of adaptive evolution
- PMID: 36223883
- PMCID: PMC10091731
- DOI: 10.1111/brv.12910
A generalised approach to the study and understanding of adaptive evolution
Abstract
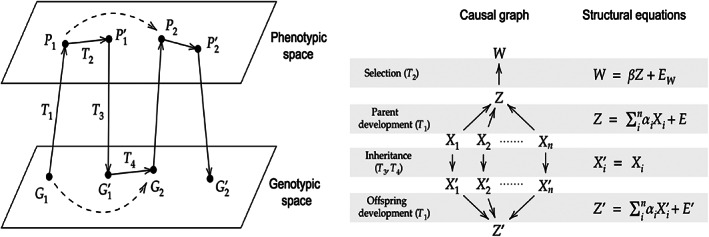
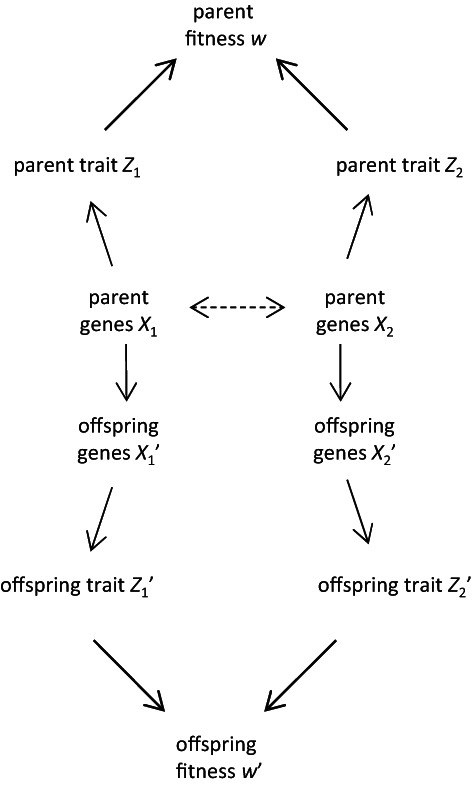
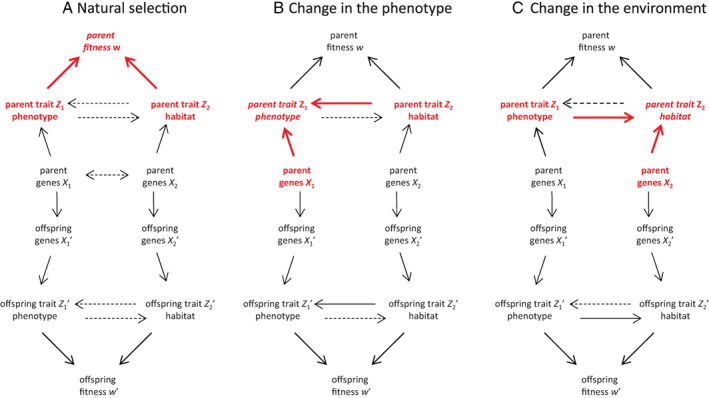
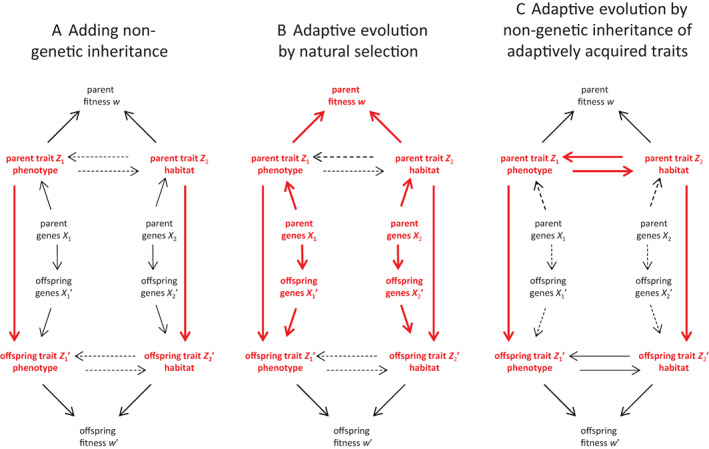
Evolutionary theory has made large impacts on our understanding and management of the world, in part because it has been able to incorporate new data and new insights successfully. Nonetheless, there is currently a tension between certain biological phenomena and mainstream evolutionary theory. For example, how does the inheritance of molecular epigenetic changes fit into mainstream evolutionary theory? Is niche construction an evolutionary process? Is local adaptation via habitat choice also adaptive evolution? These examples suggest there is scope (and perhaps even a need) to broaden our views on evolution. We identify three aspects whose incorporation into a single framework would enable a more generalised approach to the understanding and study of adaptive evolution: (i) a broadened view of extended phenotypes; (ii) that traits can respond to each other; and (iii) that inheritance can be non-genetic. We use causal modelling to integrate these three aspects with established views on the variables and mechanisms that drive and allow for adaptive evolution. Our causal model identifies natural selection and non-genetic inheritance of adaptive parental responses as two complementary yet distinct and independent drivers of adaptive evolution. Both drivers are compatible with the Price equation; specifically, non-genetic inheritance of parental responses is captured by an often-neglected component of the Price equation. Our causal model is general and simplified, but can be adjusted flexibly in terms of variables and causal connections, depending on the research question and/or biological system. By revisiting the three examples given above, we show how to use it as a heuristic tool to clarify conceptual issues and to help design empirical research. In contrast to a gene-centric view defining evolution only in terms of genetic change, our generalised approach allows us to see evolution as a change in the whole causal structure, consisting not just of genetic but also of phenotypic and environmental variables.
Keywords: Price equation; adaptation; causal modelling; evolutionary synthesis; evolutionary theory; non-genetic inheritance; plasticity.
© 2022 The Authors. Biological Reviews published by John Wiley & Sons Ltd on behalf of Cambridge Philosophical Society.
Figures
Similar articles
-
Generational stability of epigenetic transgenerational inheritance facilitates adaptation and evolution.Epigenetics. 2024 Dec;19(1):2380929. doi: 10.1080/15592294.2024.2380929. Epub 2024 Aug 5. Epigenetics. 2024. PMID: 39104183 Free PMC article. Review.
-
Niche construction and the environmental term of the price equation: How natural selection changes when organisms alter their environments.Evol Dev. 2023 Nov;25(6):451-469. doi: 10.1111/ede.12452. Epub 2023 Aug 2. Evol Dev. 2023. PMID: 37530093
-
Social competition as a driver of phenotype-environment correlations: implications for ecology and evolution.Biol Rev Camb Philos Soc. 2021 Dec;96(6):2561-2572. doi: 10.1111/brv.12768. Epub 2021 Jun 18. Biol Rev Camb Philos Soc. 2021. PMID: 34145714 Free PMC article.
-
Beyond genotype-phenotype maps: Toward a phenotype-centered perspective on evolution.Bioessays. 2022 Sep;44(9):e2100225. doi: 10.1002/bies.202100225. Epub 2022 Jul 21. Bioessays. 2022. PMID: 35863907
-
Constraints on the evolution of adaptive phenotypic plasticity in plants.New Phytol. 2005 Apr;166(1):49-60. doi: 10.1111/j.1469-8137.2004.01296.x. New Phytol. 2005. PMID: 15760350 Review.
Cited by
-
Epigenetics and seasonal timing in animals: a concise review.J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024 Jul;210(4):565-574. doi: 10.1007/s00359-023-01673-3. Epub 2023 Sep 11. J Comp Physiol A Neuroethol Sens Neural Behav Physiol. 2024. PMID: 37695537 Free PMC article. Review.
-
Comparative transcriptome and methylome of polar bears, giant and red pandas reveal diet-driven adaptive evolution.Evol Appl. 2024 Jun 17;17(6):e13731. doi: 10.1111/eva.13731. eCollection 2024 Jun. Evol Appl. 2024. PMID: 38894980 Free PMC article.
-
On Non-Random Mating, Adaptive Evolution, and Information Theory.Biology (Basel). 2024 Nov 25;13(12):970. doi: 10.3390/biology13120970. Biology (Basel). 2024. PMID: 39765637 Free PMC article.
-
Matching Habitat Choice and the Evolution of a Species' Range.Bull Math Biol. 2025 May 7;87(6):70. doi: 10.1007/s11538-025-01445-x. Bull Math Biol. 2025. PMID: 40332627 Free PMC article.
-
Resource choice during ontogeny enhances both the short- and longer-term welfare of laying hen pullets.Sci Rep. 2024 Feb 9;14(1):3360. doi: 10.1038/s41598-024-53039-7. Sci Rep. 2024. PMID: 38336837 Free PMC article.
References
-
- Adrian‐Kalchhauser, I. , Sultan, S. E. , Shama, L. N. S. , Spence‐Jones, H. , Tiso, S. , Keller Valsecchi, C. I. & Weissing, F. J. (2020). Understanding “non‐genetic” inheritance: insights from molecular‐evolutionary crosstalk. Trends in Ecology and Evolution 35, 1078–1089. - PubMed
-
- Ågren, A. J. (2021). The Gene's‐Eye View of Evolution. Oxford University Press, Oxford.
-
- Akcali, C. K. & Porter, C. K. (2017). Comment on Van Belleghem et al. 2016: Habitat choice mechanisms in speciation and other forms of diversification. Evolution 71, 2754–2761. - PubMed
-
- Anastasiadi, D. , Venney, C. J. , Bernatchez, L. & Wellenreuther, M. (2021). Epigenetic inheritance and reproductive mode in plants and animals. Trends in Ecology and Evolution 36, 1124–1140. - PubMed
-
- Araya‐Ajoy, Y. , Westneat, D. F. & Wright, J. (2020). Pathways to social evolution and their evolutionary feedbacks. Evolution 74, 1894–1907. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
