

Lineage-selective super enhancers mediate core regulatory circuitry during adipogenic and osteogenic differentiation of human mesenchymal stem cells
- PMID: 36224171
- PMCID: PMC9556616
- DOI: 10.1038/s41419-022-05309-3
Lineage-selective super enhancers mediate core regulatory circuitry during adipogenic and osteogenic differentiation of human mesenchymal stem cells
Abstract
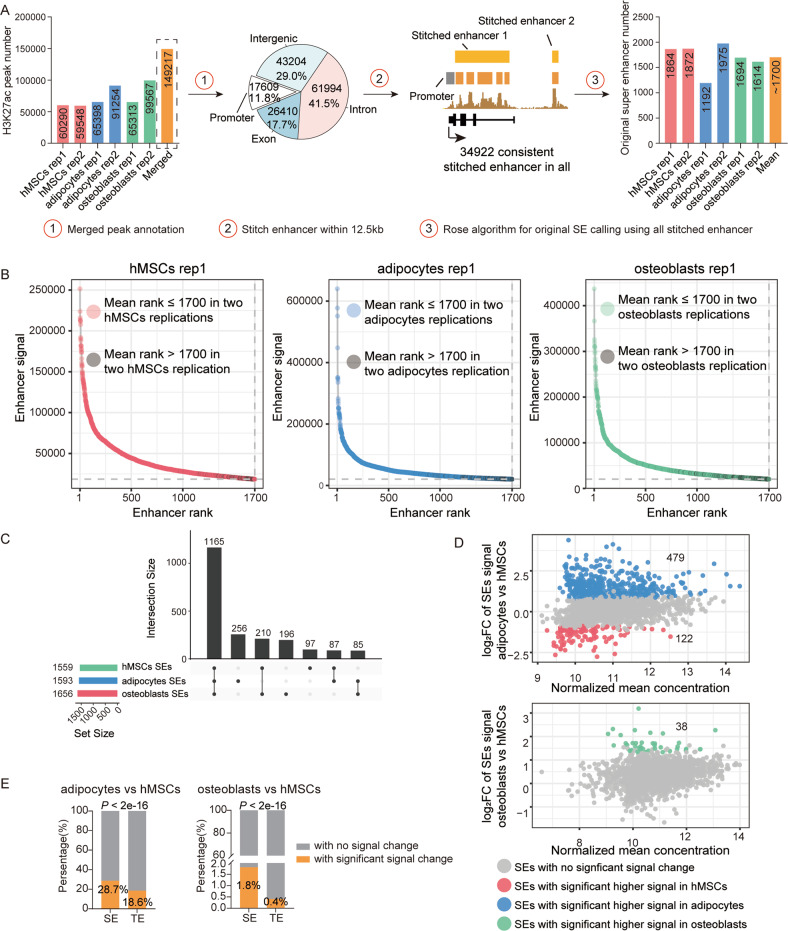
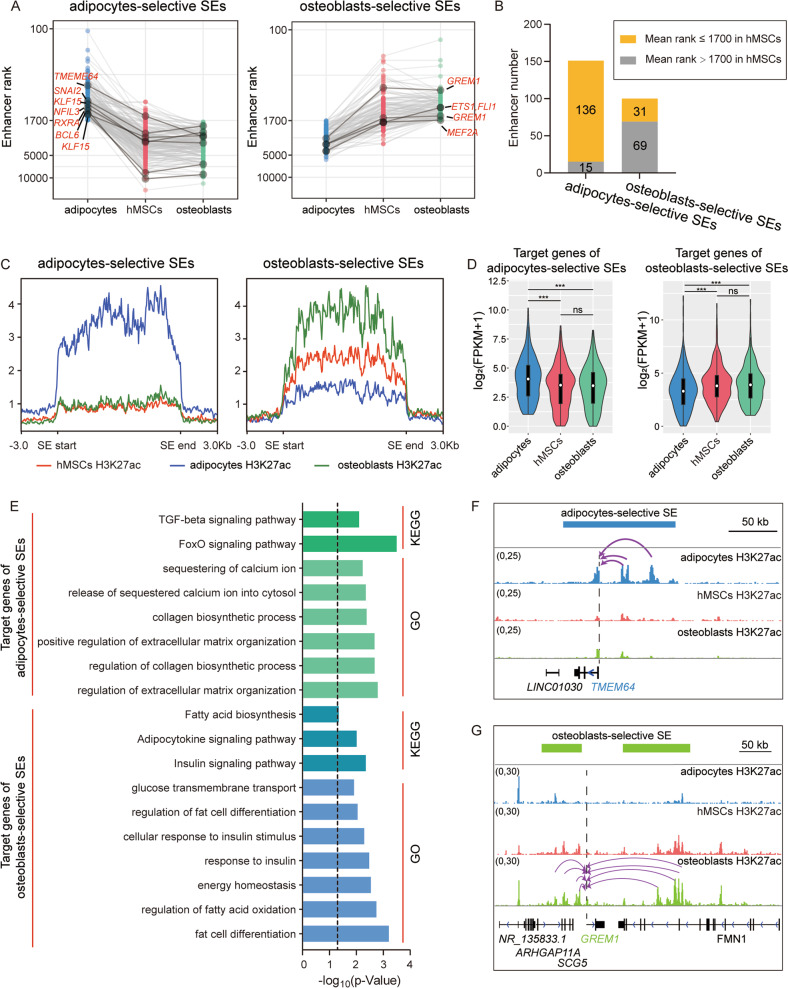
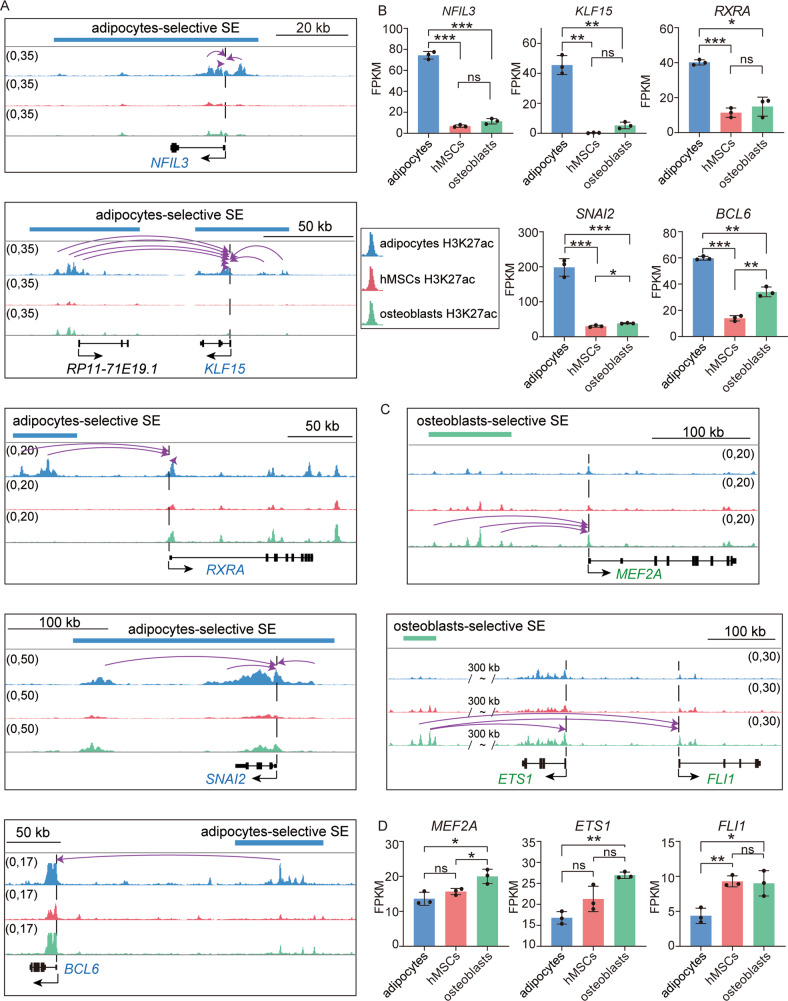
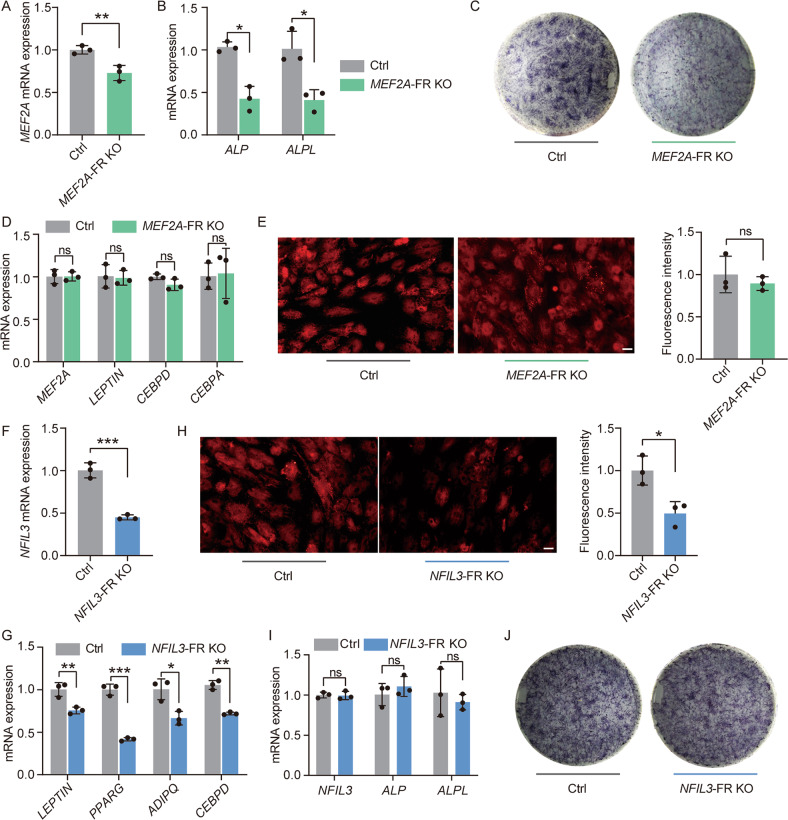
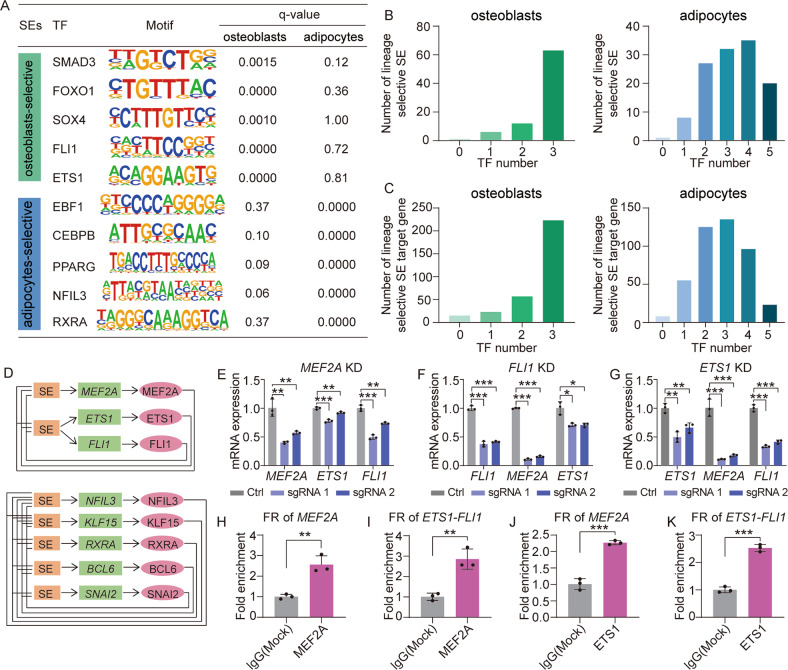
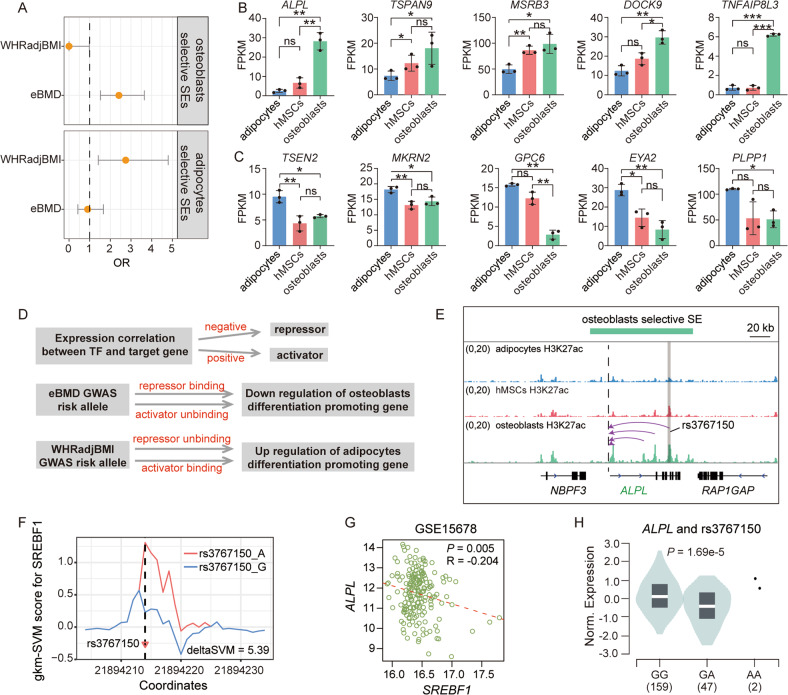
Human mesenchymal stem cells (hMSCs) can be differentiated into osteoblasts and adipocytes. During these processes, super enhancers (SEs) play important roles. Here, we performed comprehensive characterization of the SEs changes associated with adipogenic and osteogenic differentiation of hMSCs, and revealed that SEs changed more dramatically compared with typical enhancers. We identified a set of lineage-selective SEs, whose target genes were enriched with cell type-specific functions. Functional experiments in lineage-selective SEs demonstrated their specific roles in directed differentiation of hMSCs. We also found that some key transcription factors regulated by lineage-selective SEs could form core regulatory circuitry (CRC) to regulate each other's expression and control the hMSCs fate determination. In addition, we found that GWAS SNPs of osteoporosis and obesity were significantly enriched in osteoblasts-selective SEs or adipocytes-selective SEs, respectively. Taken together, our studies unveiled important roles of lineage-selective SEs in hMSCs differentiation into osteoblasts and adipocytes.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Lineage-specific rearrangement of chromatin loops and epigenomic features during adipocytes and osteoblasts commitment.Cell Death Differ. 2022 Dec;29(12):2503-2518. doi: 10.1038/s41418-022-01035-7. Epub 2022 Jul 29. Cell Death Differ. 2022. PMID: 35906483 Free PMC article.
-
Lineage-specific exosomes could override extracellular matrix mediated human mesenchymal stem cell differentiation.Biomaterials. 2018 Nov;182:312-322. doi: 10.1016/j.biomaterials.2018.08.027. Epub 2018 Aug 11. Biomaterials. 2018. PMID: 30153612 Free PMC article.
-
Downregulation of SUV39H1 and CITED2 Exerts Additive Effect on Promoting Adipogenic Commitment of Human Mesenchymal Stem Cells.Stem Cells Dev. 2021 May 1;30(9):485-501. doi: 10.1089/scd.2020.0190. Epub 2021 Apr 13. Stem Cells Dev. 2021. PMID: 33691475 Free PMC article.
-
The role of zinc finger proteins in the fate determination of mesenchymal stem cells during osteogenic and adipogenic differentiation.Int J Biochem Cell Biol. 2024 Feb;167:106507. doi: 10.1016/j.biocel.2023.106507. Epub 2023 Dec 23. Int J Biochem Cell Biol. 2024. PMID: 38142772 Review.
-
The role of microRNAs in cell fate determination of mesenchymal stem cells: balancing adipogenesis and osteogenesis.BMB Rep. 2015 Jun;48(6):319-23. doi: 10.5483/bmbrep.2015.48.6.206. BMB Rep. 2015. PMID: 25341923 Free PMC article. Review.
Cited by
-
Experimental Validation and Prediction of Super-Enhancers: Advances and Challenges.Cells. 2023 Apr 19;12(8):1191. doi: 10.3390/cells12081191. Cells. 2023. PMID: 37190100 Free PMC article. Review.
-
Obesity and lipid metabolism in the development of osteoporosis (Review).Int J Mol Med. 2024 Jul;54(1):61. doi: 10.3892/ijmm.2024.5385. Epub 2024 May 31. Int J Mol Med. 2024. PMID: 38818830 Free PMC article. Review.
-
The role of the haematopoietic stem cell niche in development and ageing.Nat Rev Mol Cell Biol. 2025 Jan;26(1):32-50. doi: 10.1038/s41580-024-00770-8. Epub 2024 Sep 10. Nat Rev Mol Cell Biol. 2025. PMID: 39256623 Review.
-
Genetic transcriptional regulation profiling of cartilage reveals pathogenesis of osteoarthritis.EBioMedicine. 2025 Jul;117:105821. doi: 10.1016/j.ebiom.2025.105821. Epub 2025 Jun 26. EBioMedicine. 2025. PMID: 40577937 Free PMC article.
References
-
- Peister A, Mellad JA, Larson BL, Hall BM, Gibson LF, Prockop DJ. Adult stem cells from bone marrow (MSCs) isolated from different strains of inbred mice vary in surface epitopes, rates of proliferation, and differentiation potential. Blood. 2004;103:1662–8. doi: 10.1182/blood-2003-09-3070. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
