

Generation and characterization of human U-2 OS cell lines with the CRISPR/Cas9-edited protoporphyrinogen oxidase IX gene
- PMID: 36224252
- PMCID: PMC9556554
- DOI: 10.1038/s41598-022-21147-x
Generation and characterization of human U-2 OS cell lines with the CRISPR/Cas9-edited protoporphyrinogen oxidase IX gene
Abstract
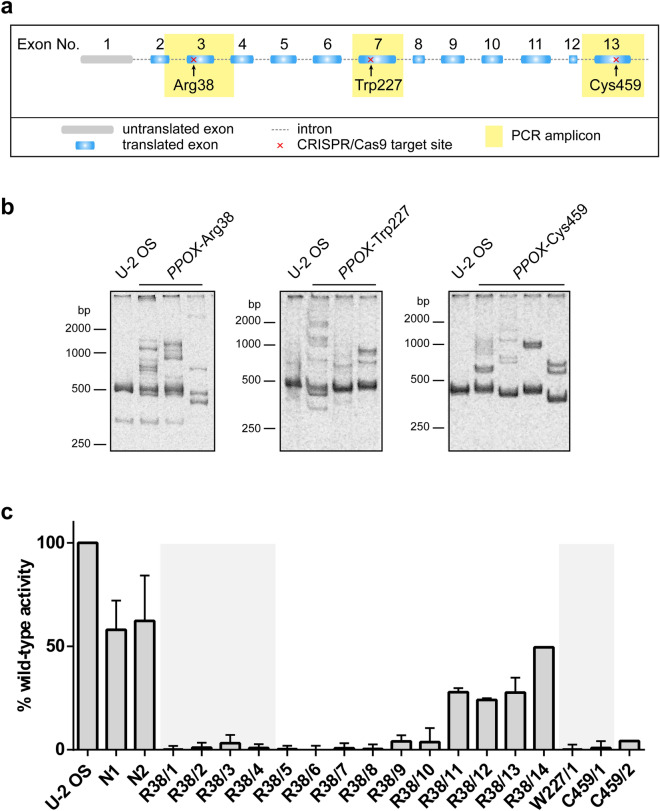
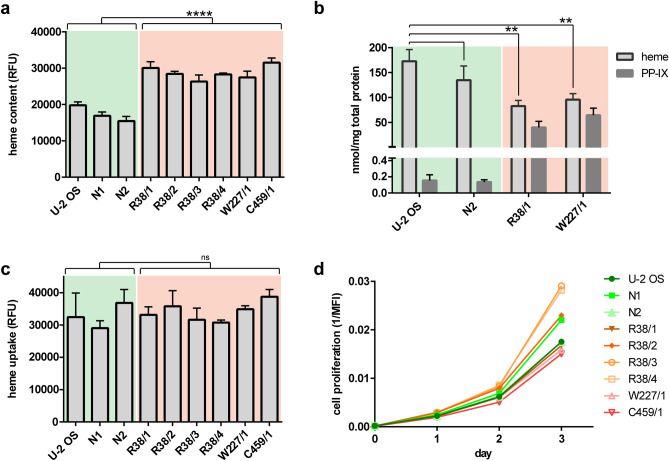
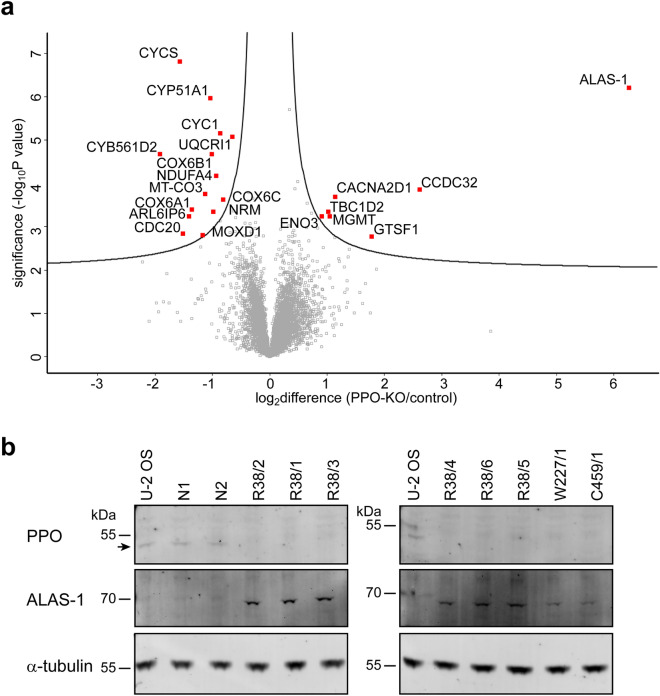
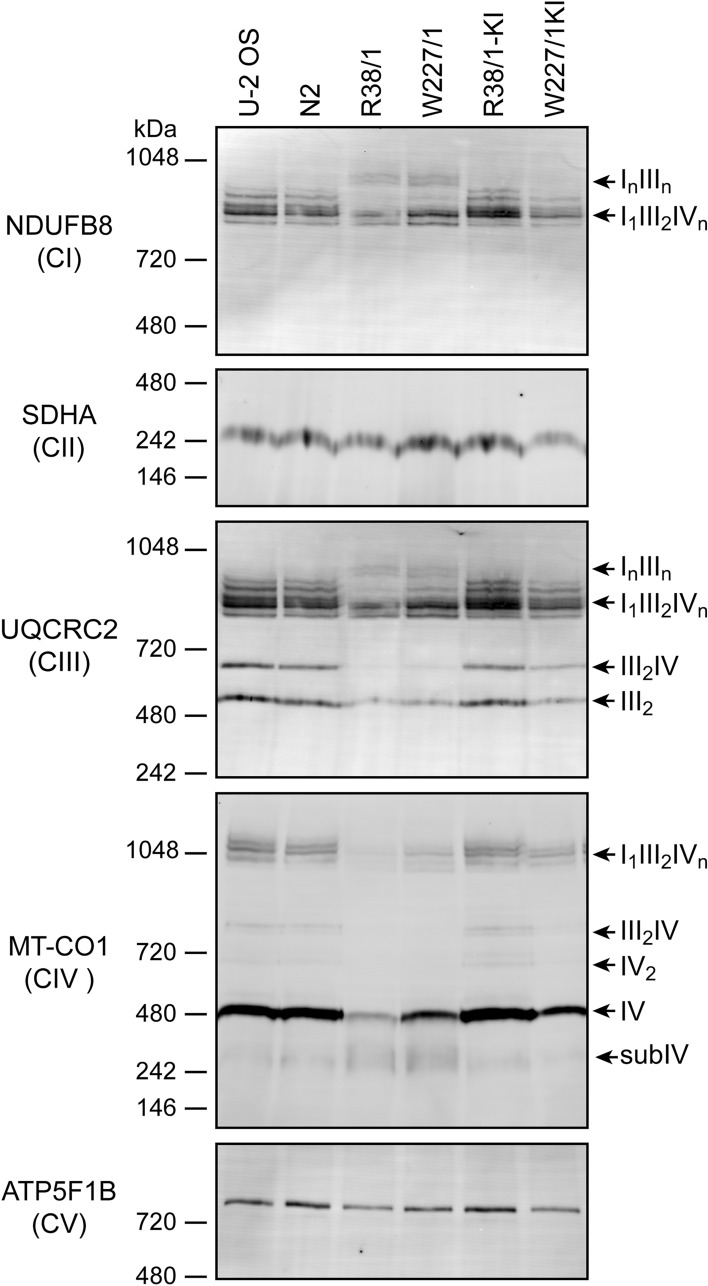
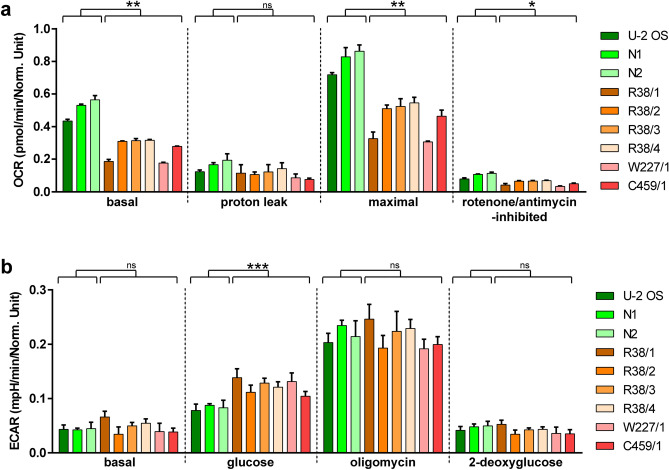
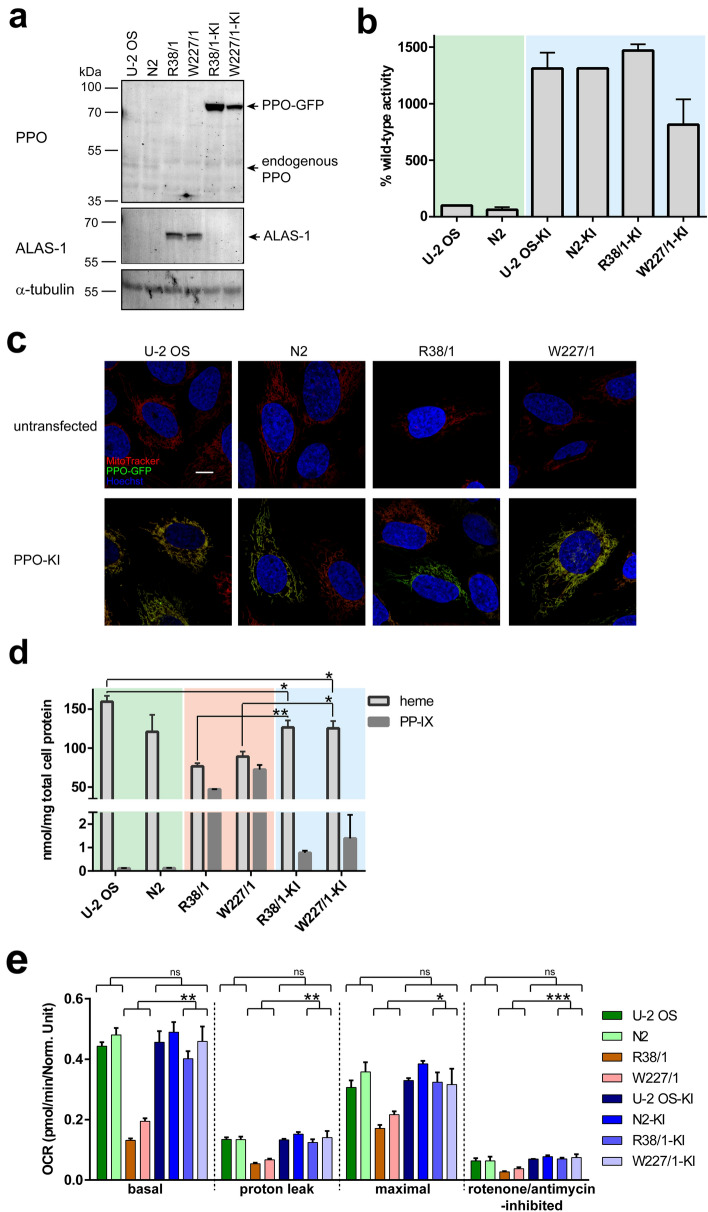
In humans, disruptions in the heme biosynthetic pathway are associated with various types of porphyrias, including variegate porphyria that results from the decreased activity of protoporphyrinogen oxidase IX (PPO; E.C.1.3.3.4), the enzyme catalyzing the penultimate step of the heme biosynthesis. Here we report the generation and characterization of human cell lines, in which PPO was inactivated using the CRISPR/Cas9 system. The PPO knock-out (PPO-KO) cell lines are viable with the normal proliferation rate and show massive accumulation of protoporphyrinogen IX, the PPO substrate. Observed low heme levels trigger a decrease in the amount of functional heme containing respiratory complexes III and IV and overall reduced oxygen consumption rates. Untargeted proteomics further revealed dysregulation of 22 cellular proteins, including strong upregulation of 5-aminolevulinic acid synthase, the major regulatory protein of the heme biosynthesis, as well as additional ten targets with unknown association to heme metabolism. Importantly, knock-in of PPO into PPO-KO cells rescued their wild-type phenotype, confirming the specificity of our model. Overall, our model system exploiting a non-erythroid human U-2 OS cell line reveals physiological consequences of the PPO ablation at the cellular level and can serve as a tool to study various aspects of dysregulated heme metabolism associated with variegate porphyria.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
Similar articles
-
Characteristics of human protoporphyrinogen oxidase in controls and variegate porphyrias.Cell Mol Biol (Noisy-le-grand). 1997 Feb;43(1):67-73. Cell Mol Biol (Noisy-le-grand). 1997. PMID: 9074790
-
The cyanobacterial protoporphyrinogen oxidase HemJ is a new b-type heme protein functionally coupled with coproporphyrinogen III oxidase.J Biol Chem. 2018 Aug 10;293(32):12394-12404. doi: 10.1074/jbc.RA118.003441. Epub 2018 Jun 20. J Biol Chem. 2018. PMID: 29925590 Free PMC article.
-
Protoporphyrin accumulation by mitogen stimulated lymphocytes and protoporphyrinogen oxidase activity in patients with porphyria variegata and erythropoietic protoporphyria: evidence for deficiency of protoporphyrinogen oxidase and ferrochelatase in both diseases.Br J Haematol. 1985 May;60(1):65-74. doi: 10.1111/j.1365-2141.1985.tb07386.x. Br J Haematol. 1985. PMID: 3924091
-
Variegate porphyria: identification of a nonsense mutation in the protoporphyrinogen oxidase gene.J Invest Dermatol. 1998 Apr;110(4):449-51. doi: 10.1046/j.1523-1747.1998.00147.x. J Invest Dermatol. 1998. PMID: 9540990 Review.
-
Homozygous variegate porphyria: identification of mutations on both alleles of the protoporphyrinogen oxidase gene in a severely affected proband.J Invest Dermatol. 1998 Apr;110(4):452-5. doi: 10.1046/j.1523-1747.1998.00148.x. J Invest Dermatol. 1998. PMID: 9540991 Review.
Cited by
-
Recent advances in therapeutic CRISPR-Cas9 genome editing: mechanisms and applications.Mol Biomed. 2023 Apr 7;4(1):10. doi: 10.1186/s43556-023-00115-5. Mol Biomed. 2023. PMID: 37027099 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
