

Role of glycosphingolipid SSEA-3 and FGF2 in the stemness and lineage commitment of multilineage differentiating stress enduring (MUSE) cells
- PMID: 36225120
- PMCID: PMC9816924
- DOI: 10.1111/cpr.13345
Role of glycosphingolipid SSEA-3 and FGF2 in the stemness and lineage commitment of multilineage differentiating stress enduring (MUSE) cells
Abstract
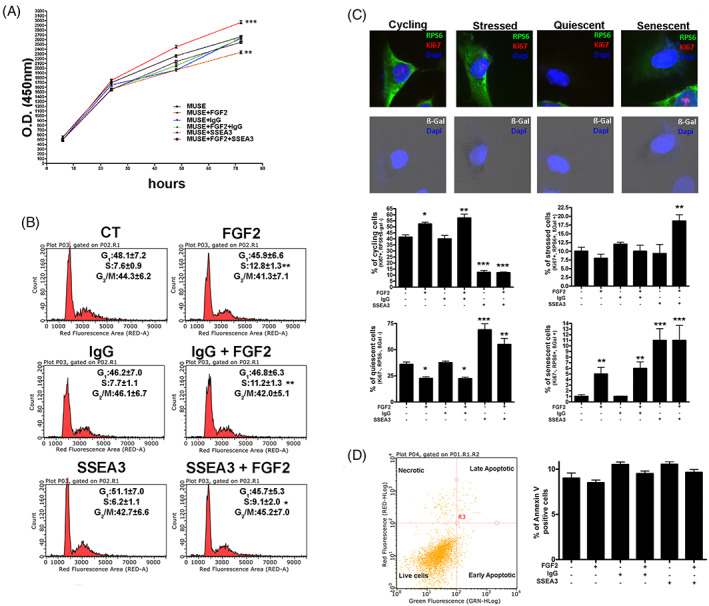
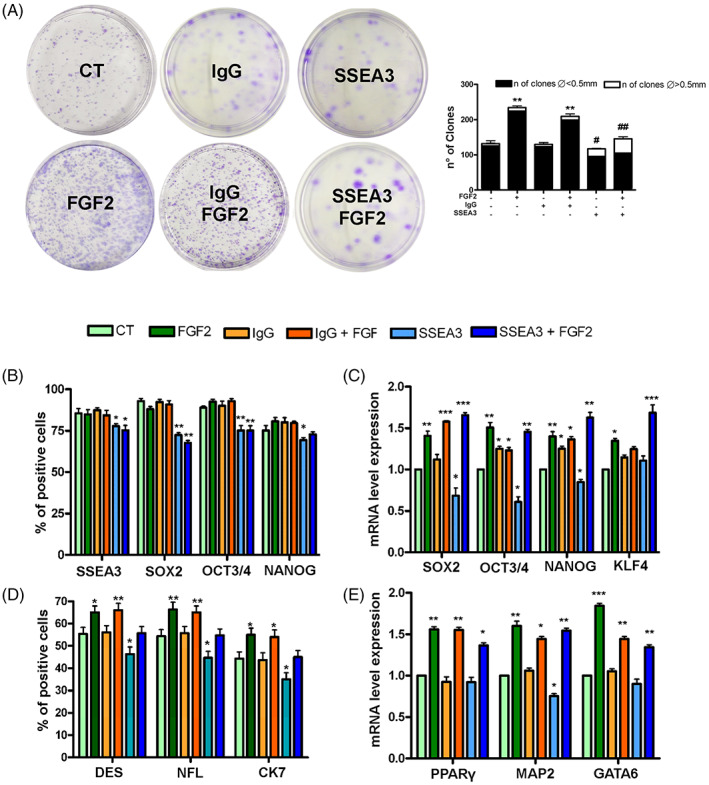
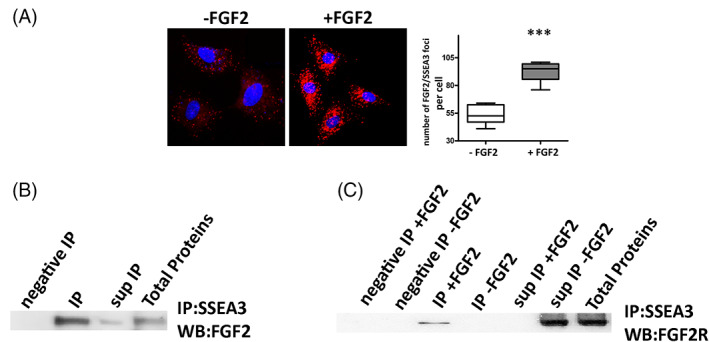
Objectives: Multilineage differentiating Stress Enduring (MUSE) cells are endogenous, stress-resistant stem cells, expressing pluripotency master genes and able to differentiate in cells of the three embryonic sheets. Stage-Specific Embryonic Antigen 3 (SSEA-3), a glycosphingolipid (GSL), is the marker for identifying MUSE cells and is used to isolate this population from mesenchymal stromal cells. GSLs modulate signal transduction by interacting with plasma membrane components. The growth factor FGF2, important for MUSE cells biology, may interact with GSLs. Specific cell surface markers represent an invaluable tool for stem cell isolation. Nonetheless their role, if any, in stem cell biology is poorly investigated. Functions of stem cells, however, depend on niche external cues, which reach cells through surface markers. We addressed the role of SSEA-3 in MUSE cell behaviour, trying to define whether SSEA-3 is just a marker or if it plays a functional role in this cell population by determining if it has any relationship with FGF2 activity.
Results: We evidenced how the SSEA-3 and FGF2 cooperation affected the self-renewal and clonogenic capacity of MUSE cells. The block of SSEA-3 significantly reduced the multilineage potential of MUSE cells with production of nullipotent clones.
Conclusions: We contributed to dissecting the mechanisms underlying MUSE cell properties for establishing successful stem-cell-based therapies and the promotion of MUSE cells as a tool for the in vitro disease model.
© 2022 The Authors. Cell Proliferation published by Beijing Institute for Stem Cell and Regenerative Medicine and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare that no conflict of interests exists.
Figures




References
-
- Yamauchi T, Yamasaki K, Tsuchiyama K, Koike S, Aiba S. A quantitative analysis of multilineage‐differentiating stress‐enduring (Muse) cells in human adipose tissue and efficacy of melanocytes induction. J Dermatol Sci. 2017;86(3):198‐205. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
