

Antibacterial T6SS effectors with a VRR-Nuc domain are structure-specific nucleases
- PMID: 36226828
- PMCID: PMC9635880
- DOI: 10.7554/eLife.82437
Antibacterial T6SS effectors with a VRR-Nuc domain are structure-specific nucleases
Abstract
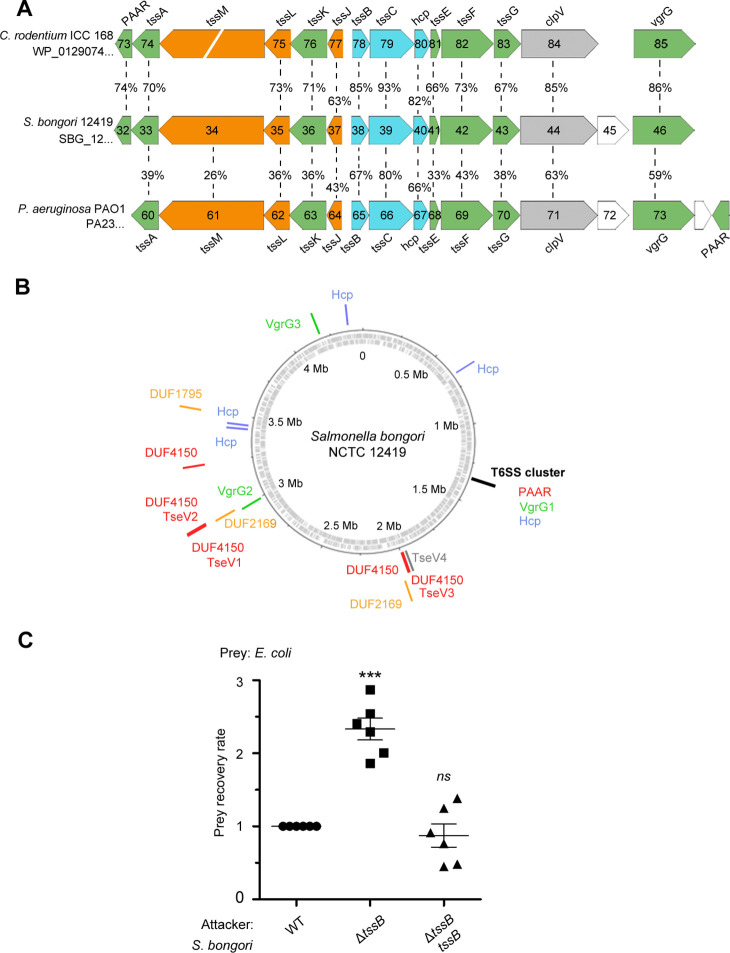
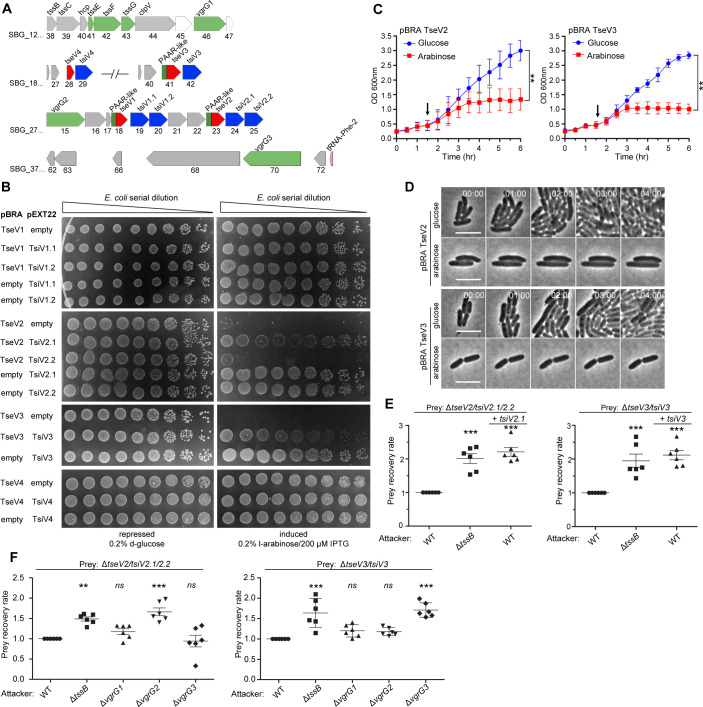
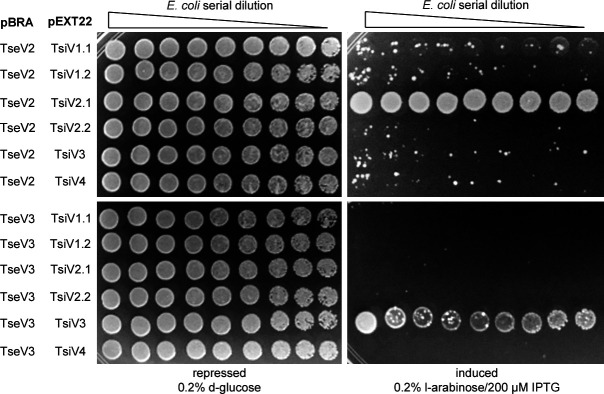
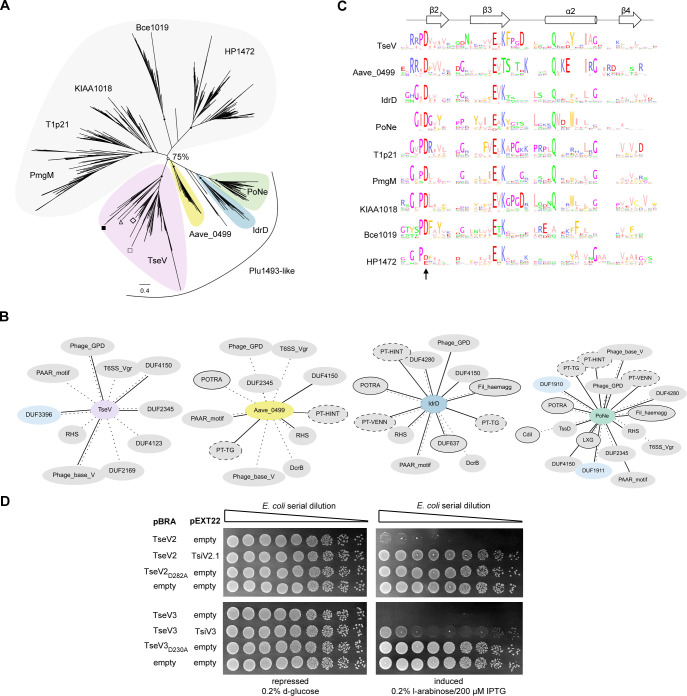
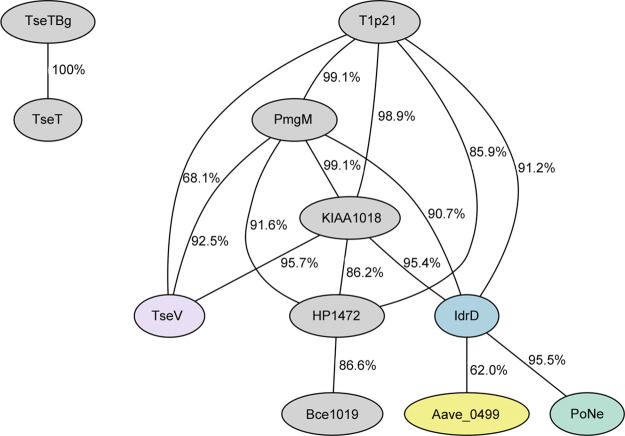
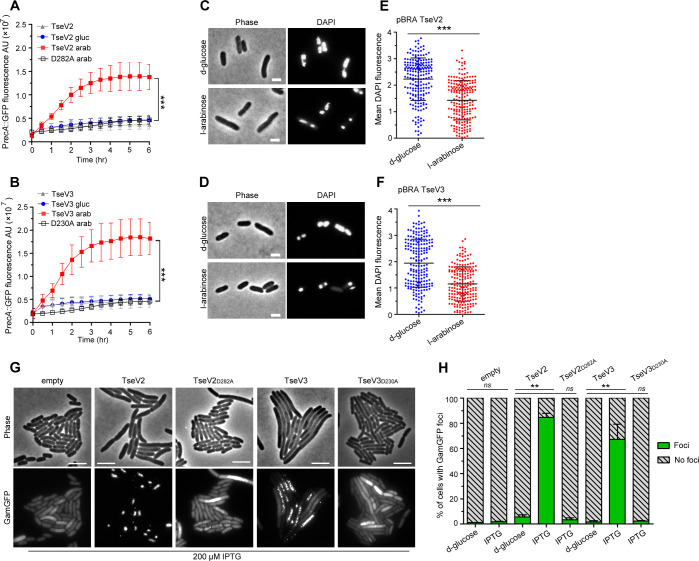
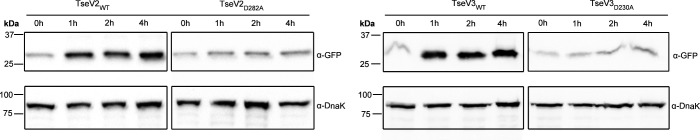
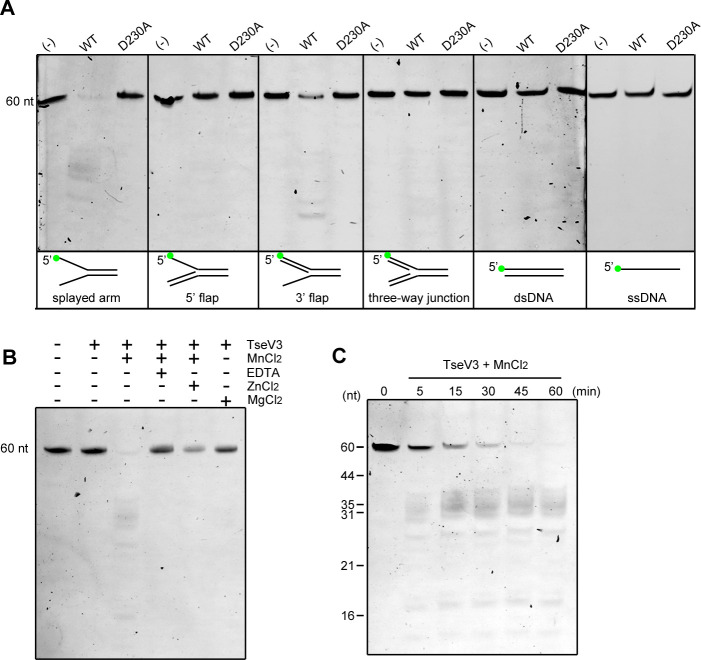
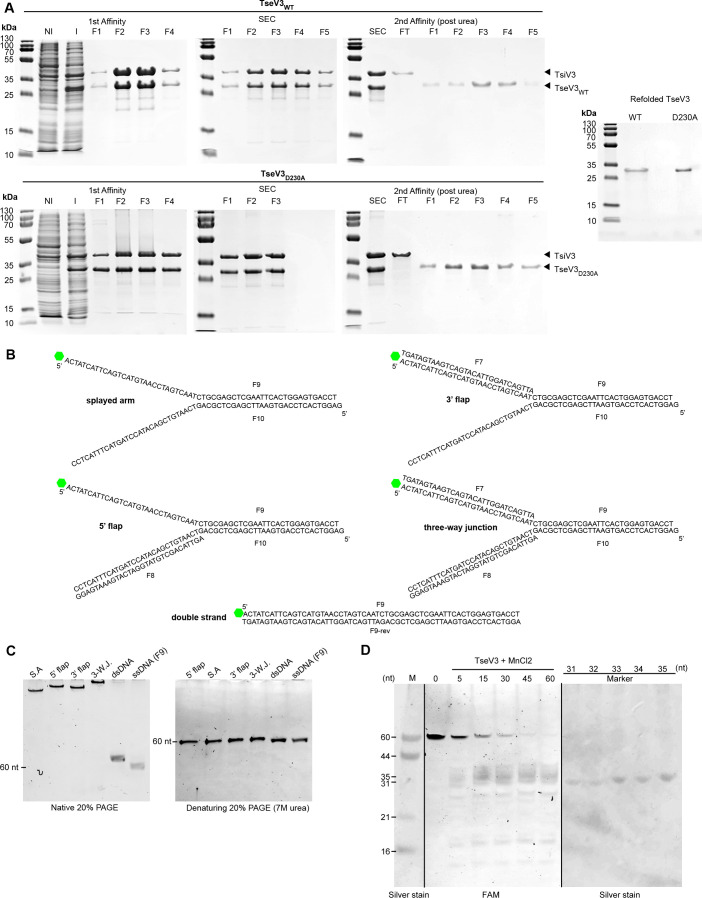
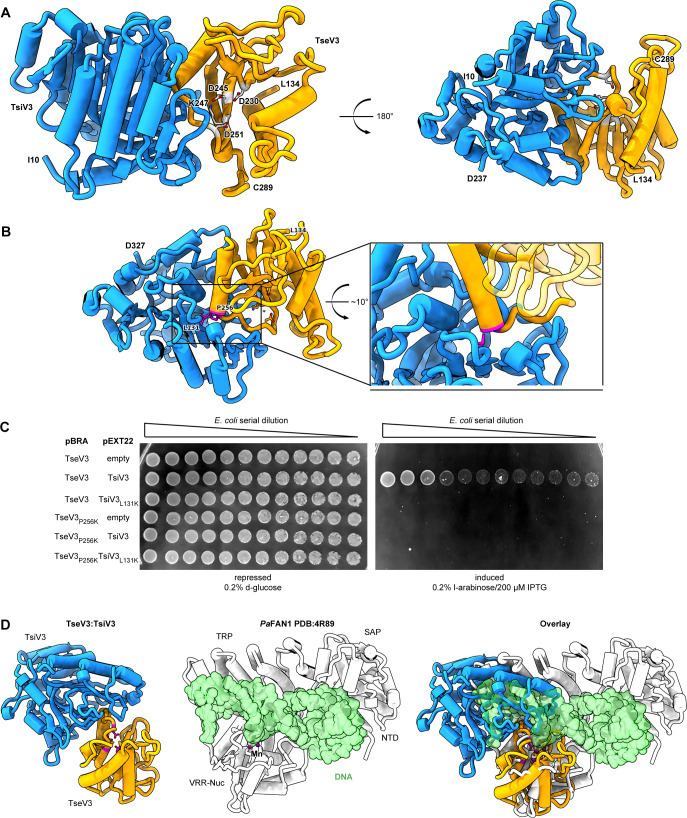
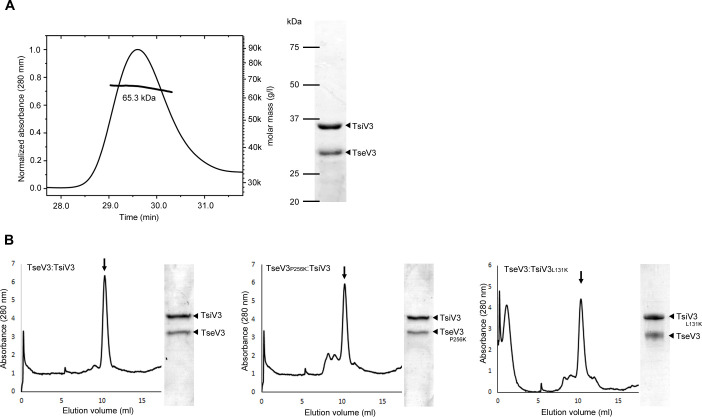
The type VI secretion system (T6SS) secretes antibacterial effectors into target competitors. Salmonella spp. encode five phylogenetically distinct T6SSs. Here, we characterize the function of the SPI-22 T6SS of Salmonella bongori showing that it has antibacterial activity and identify a group of antibacterial T6SS effectors (TseV1-4) containing an N-terminal PAAR-like domain and a C-terminal VRR-Nuc domain encoded next to cognate immunity proteins with a DUF3396 domain (TsiV1-4). TseV2 and TseV3 are toxic when expressed in Escherichia coli and bacterial competition assays confirm that TseV2 and TseV3 are secreted by the SPI-22 T6SS. Phylogenetic analysis reveals that TseV1-4 are evolutionarily related to enzymes involved in DNA repair. TseV3 recognizes specific DNA structures and preferentially cleave splayed arms, generating DNA double-strand breaks and inducing the SOS response in target cells. The crystal structure of the TseV3:TsiV3 complex reveals that the immunity protein likely blocks the effector interaction with the DNA substrate. These results expand our knowledge on the function of Salmonella pathogenicity islands, the evolution of toxins used in biological conflicts, and the endogenous mechanisms regulating the activity of these toxins.
Keywords: Salmonella; T6SS; VRR-Nuc; effector; infectious disease; microbiology; nuclease; toxin.
© 2022, Hespanhol, Sanchez-Limache et al.
Conflict of interest statement
JH, DS, GN, LM, EL, GC, CF, Rd, RG, AL, EB No competing interests declared
Figures
References
-
- Bayer-Santos E, Cenens W, Matsuyama BY, Oka GU, Di Sessa G, Mininel IDV, Alves TL, Farah CS. The opportunistic pathogen stenotrophomonas maltophilia utilizes a type IV secretion system for interbacterial killing. PLOS Pathogens. 2019;15:e1007651. doi: 10.1371/journal.ppat.1007651. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
