

In vitro neurons learn and exhibit sentience when embodied in a simulated game-world
- PMID: 36228614
- PMCID: PMC9747182
- DOI: 10.1016/j.neuron.2022.09.001
In vitro neurons learn and exhibit sentience when embodied in a simulated game-world
Abstract
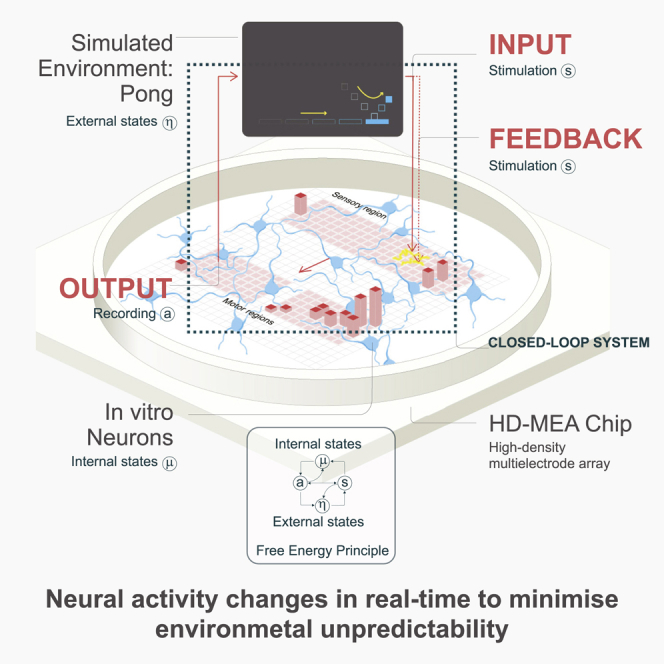
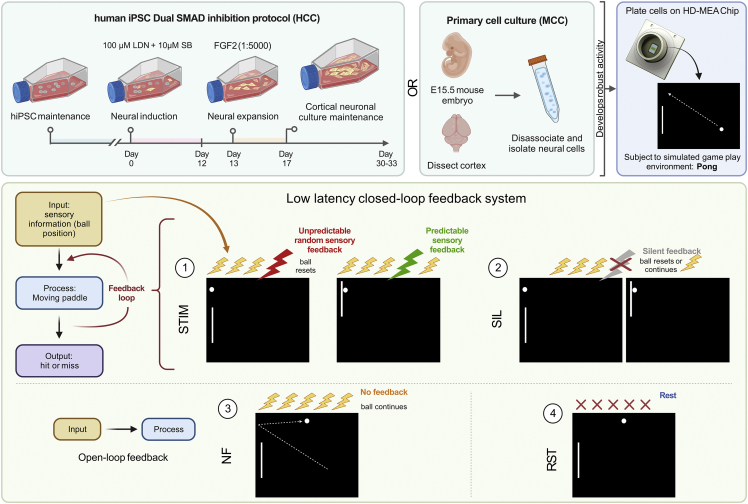
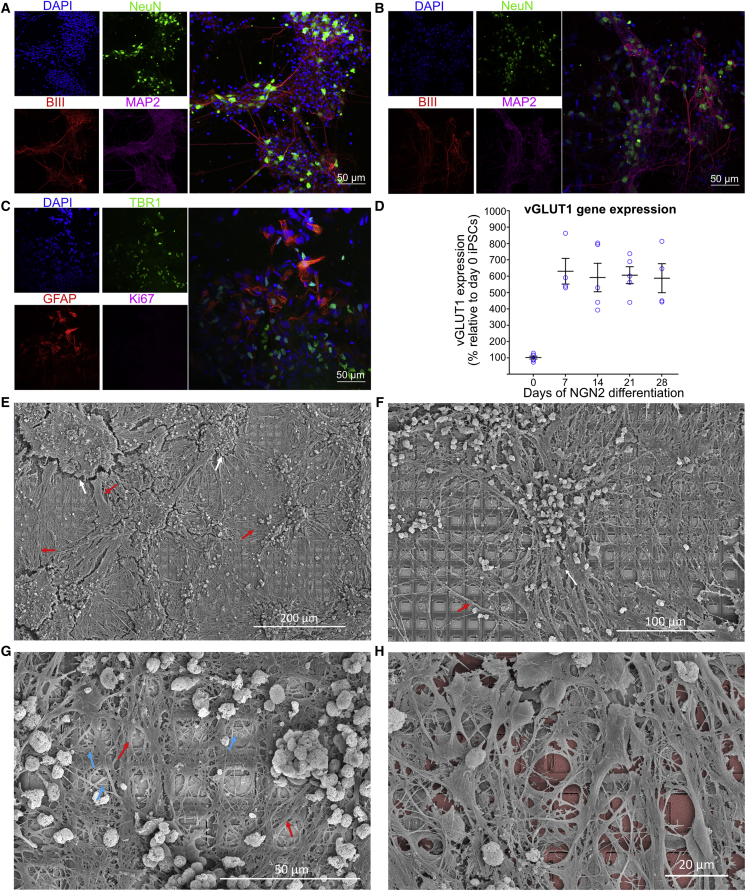
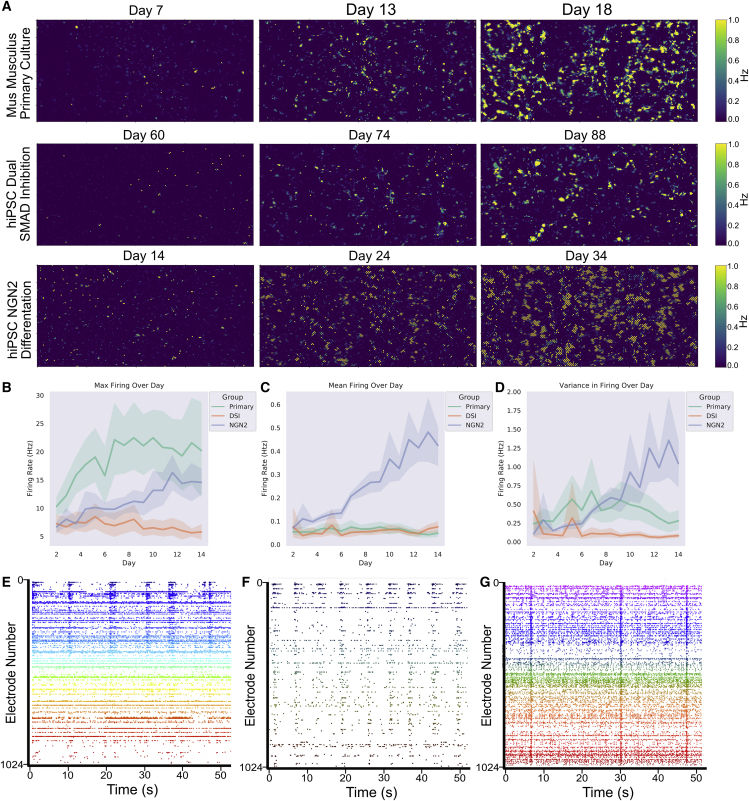
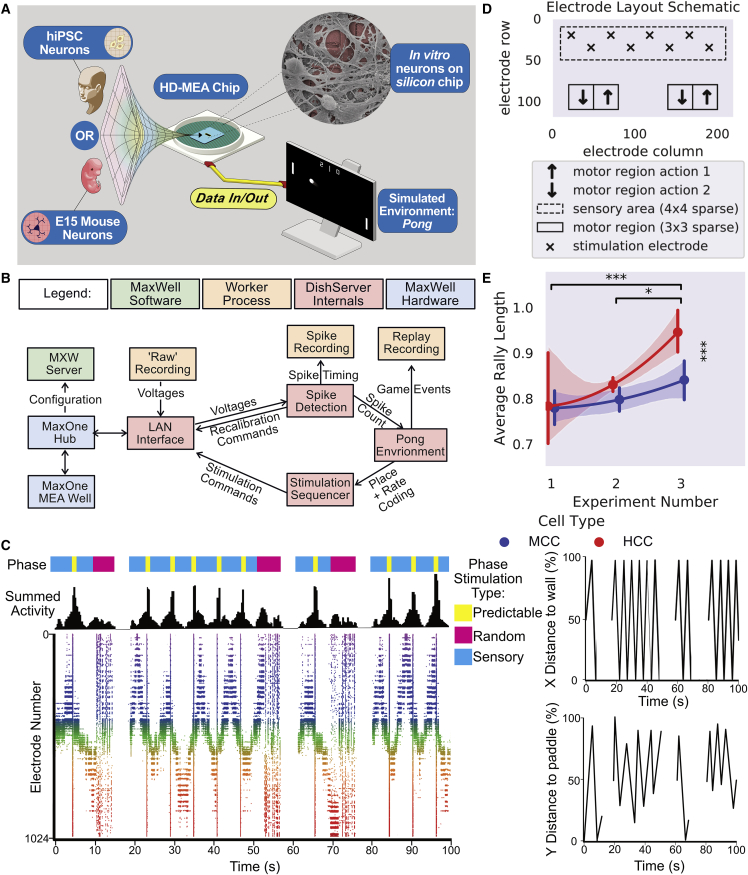
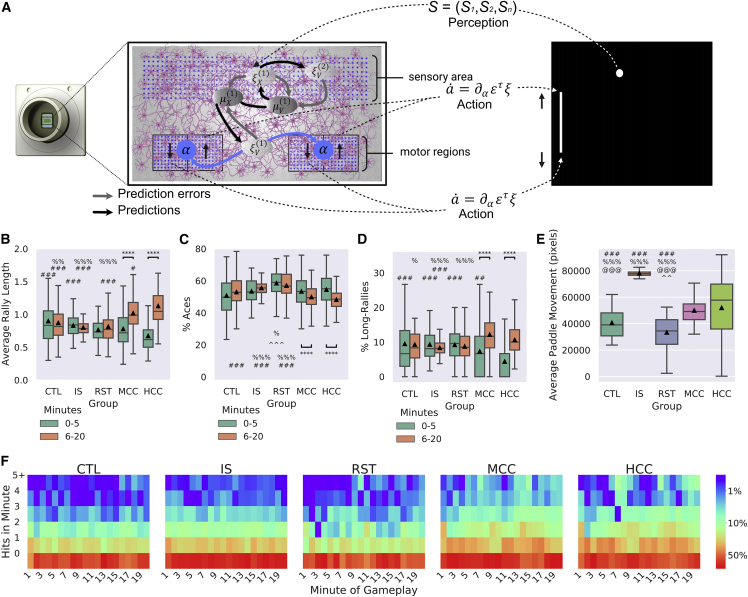
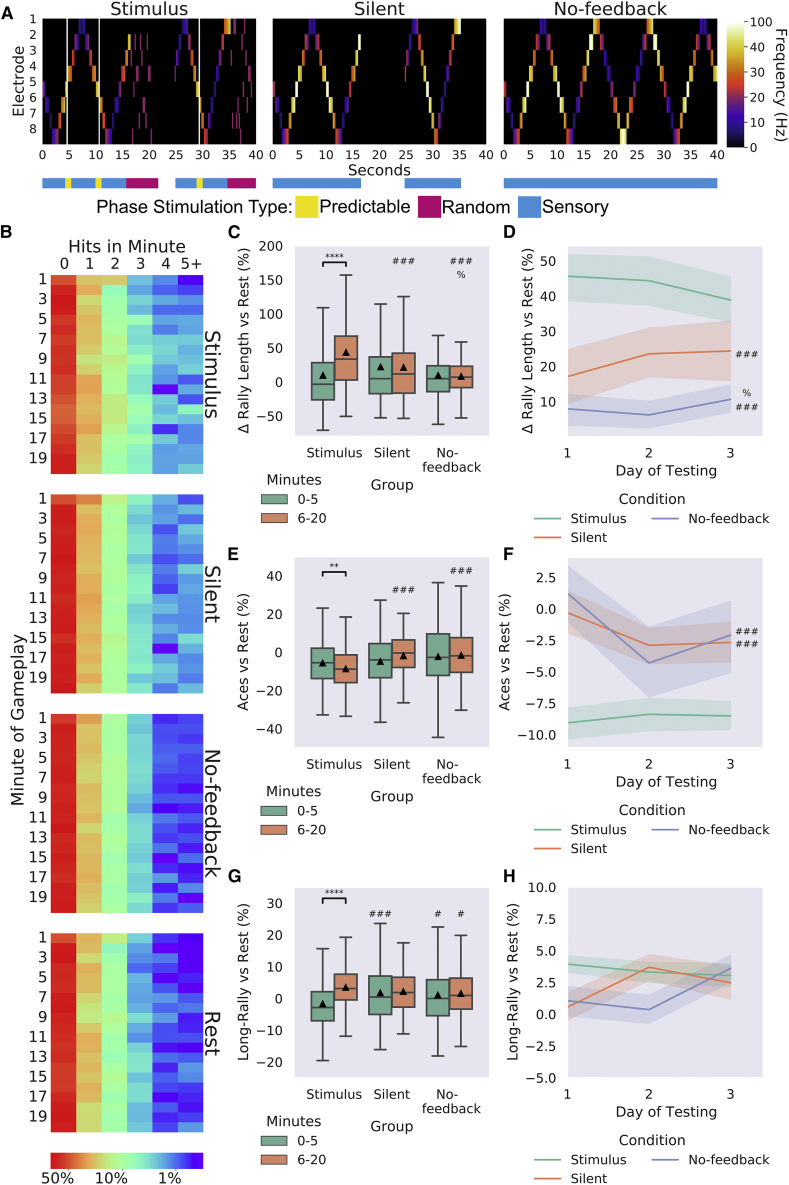
Integrating neurons into digital systems may enable performance infeasible with silicon alone. Here, we develop DishBrain, a system that harnesses the inherent adaptive computation of neurons in a structured environment. In vitro neural networks from human or rodent origins are integrated with in silico computing via a high-density multielectrode array. Through electrophysiological stimulation and recording, cultures are embedded in a simulated game-world, mimicking the arcade game "Pong." Applying implications from the theory of active inference via the free energy principle, we find apparent learning within five minutes of real-time gameplay not observed in control conditions. Further experiments demonstrate the importance of closed-loop structured feedback in eliciting learning over time. Cultures display the ability to self-organize activity in a goal-directed manner in response to sparse sensory information about the consequences of their actions, which we term synthetic biological intelligence. Future applications may provide further insights into the cellular correlates of intelligence.
Keywords: cell culture; electrophysiology; free energy principle; intelligence; in vitro; learning; microphysiological systems; neurocomputation; neurons; synthetic biological intelligence.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests B.J.K. is an employee of Cortical Labs. B.J.K. and A.C.K. are shareholders of Cortical Labs. B.J.K. and A.C.K. hold an interest in patents related to this publication. F.H. and M.K. received funding from Cortical Labs for work related to this publication.
Figures


Comment in
-
Neurons in a dish learn to play Pong - what's next?Nature. 2022 Oct;610(7932):433. doi: 10.1038/d41586-022-03229-y. Nature. 2022. PMID: 36224373 No abstract available.
-
Neuronal cultures playing Pong: First steps toward advanced screening and biological computing.Neuron. 2022 Dec 7;110(23):3855-3856. doi: 10.1016/j.neuron.2022.11.010. Neuron. 2022. PMID: 36480938
References
LinkOut - more resources
Full Text Sources
Other Literature Sources
