

Trichoderma atroviride hyphal regeneration and conidiation depend on cell-signaling processes regulated by a microRNA-like RNA
- PMID: 36239595
- PMCID: PMC9676055
- DOI: 10.1099/mgen.0.000869
Trichoderma atroviride hyphal regeneration and conidiation depend on cell-signaling processes regulated by a microRNA-like RNA
Abstract
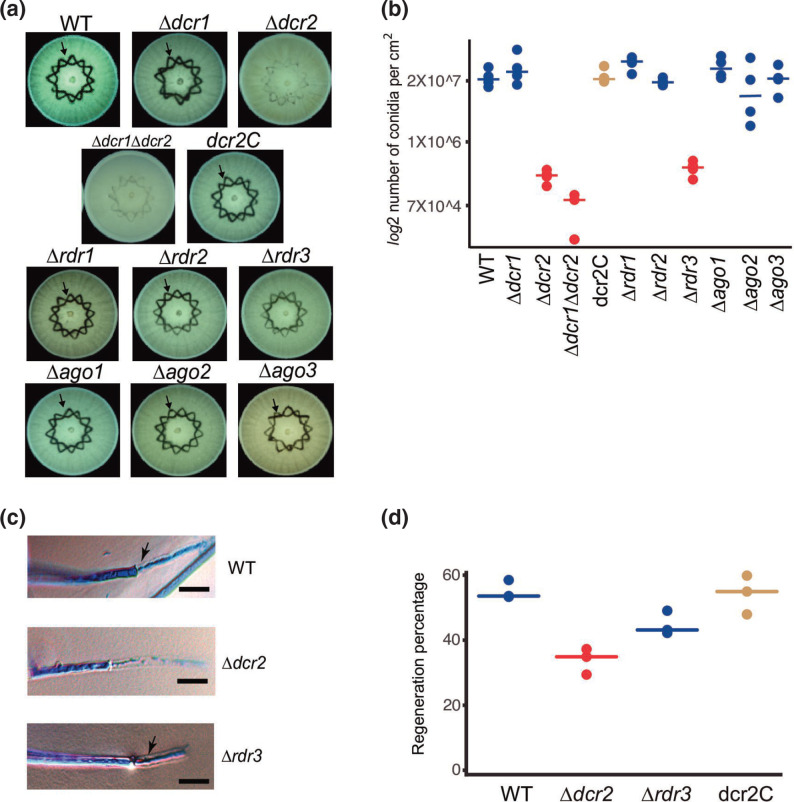
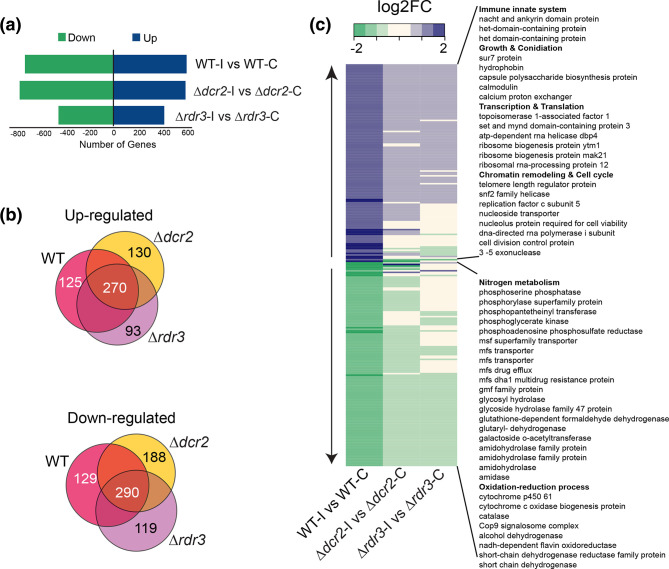
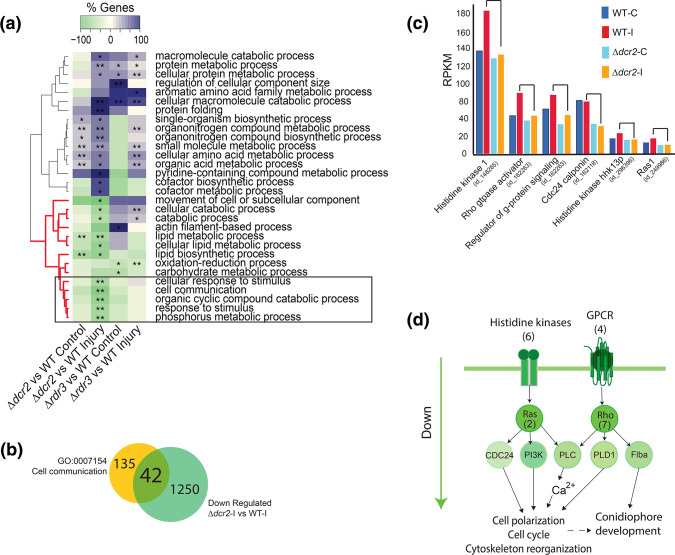
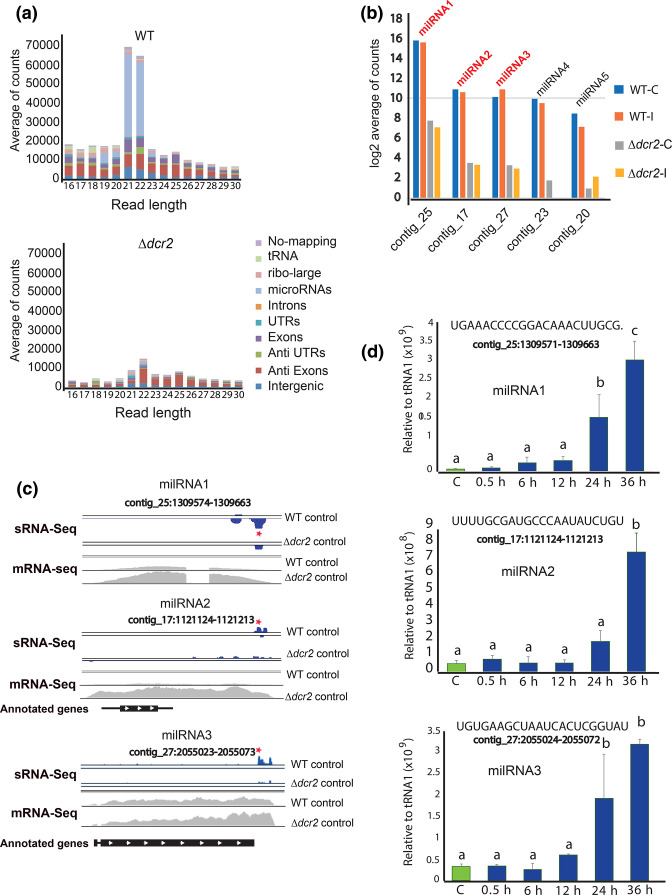
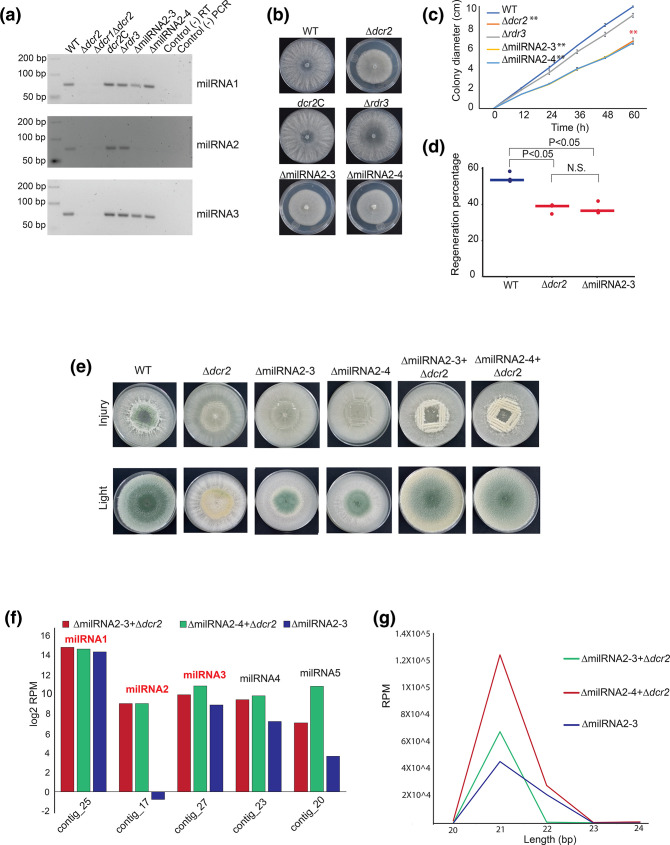
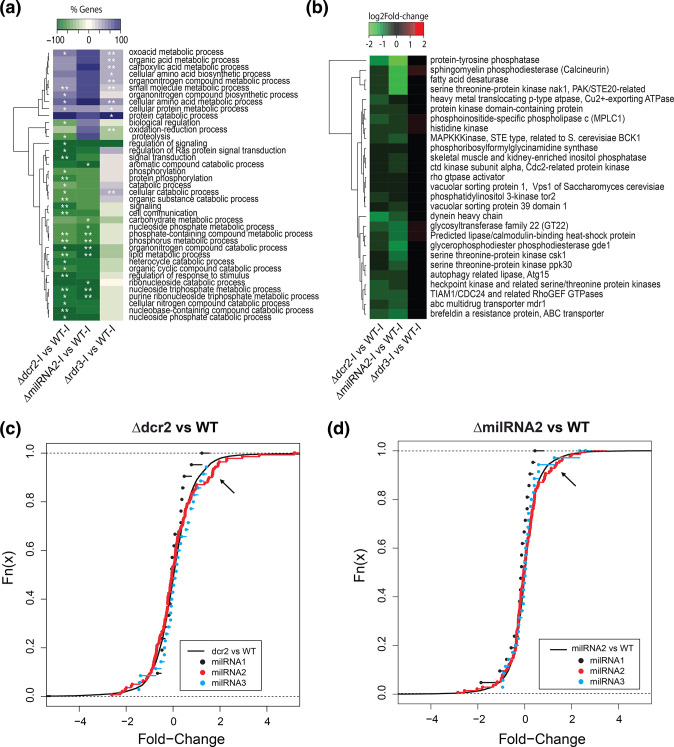
The ability to respond to injury is essential for the survival of an organism and involves analogous mechanisms in animals and plants. Such mechanisms integrate coordinated genetic and metabolic reprogramming events requiring regulation by small RNAs for adequate healing of the wounded area. We have previously reported that the response to injury of the filamentous fungus Trichoderma atroviride involves molecular mechanisms closely resembling those of plants and animals that lead to the formation of new hyphae (regeneration) and the development of asexual reproduction structures (conidiophores). However, the involvement of microRNAs in this process has not been investigated in fungi. In this work, we explore the participation of microRNA-like RNAs (milRNAs) molecules by sequencing messenger and small RNAs during the injury response of the WT strain and RNAi mutants. We found that Dcr2 appears to play an important role in hyphal regeneration and is required to produce the majority of sRNAs in T. atroviride. We also determined that the three main milRNAs produced via Dcr2 are induced during the damage-triggered developmental process. Importantly, elimination of a single milRNA phenocopied the main defects observed in the dcr2 mutant. Our results demonstrate the essential role of milRNAs in hyphal regeneration and asexual development by post-transcriptionally regulating cellular signalling processes involving phosphorylation events. These observations allow us to conclude that fungi, like plants and animals, in response to damage activate fine-tuning regulatory mechanisms.
Keywords: RNA-seq; RNAi machinery; conidiation; filamentous fungi; hyphal regeneration; milRNAs; signaling; small RNAs; transcriptome.
Conflict of interest statement
The authors declare that they have no conflicts of interests.
Figures
References
Publication types
MeSH terms
Substances
Supplementary concepts
LinkOut - more resources
Full Text Sources
Molecular Biology Databases