

Germline immortality relies on TRIM32-mediated turnover of a maternal mRNA activator in C. elegans
- PMID: 36240265
- PMCID: PMC9565796
- DOI: 10.1126/sciadv.abn0897
Germline immortality relies on TRIM32-mediated turnover of a maternal mRNA activator in C. elegans
Abstract
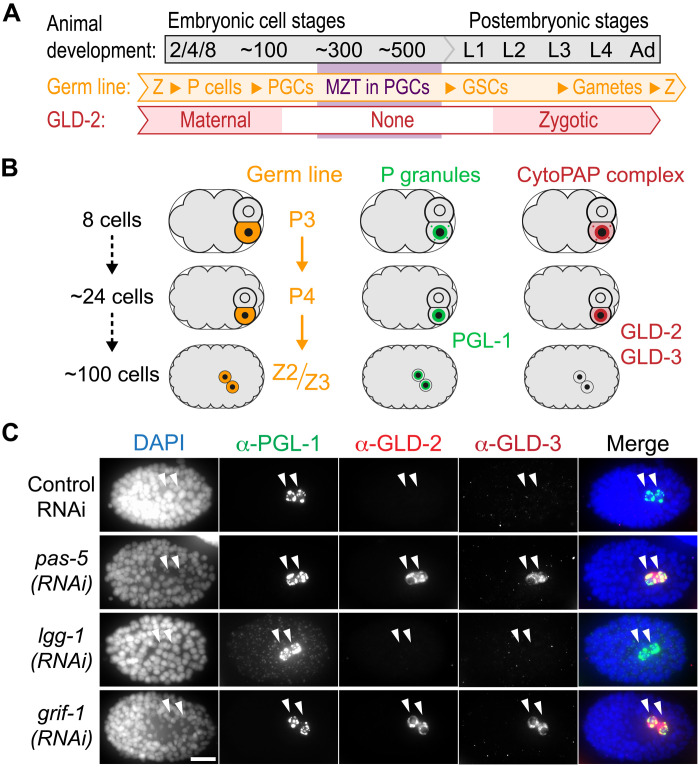
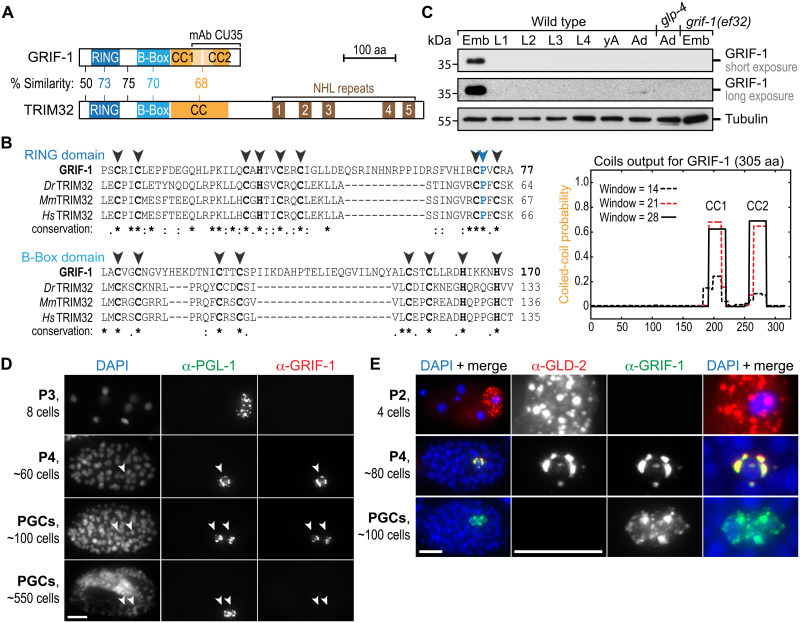
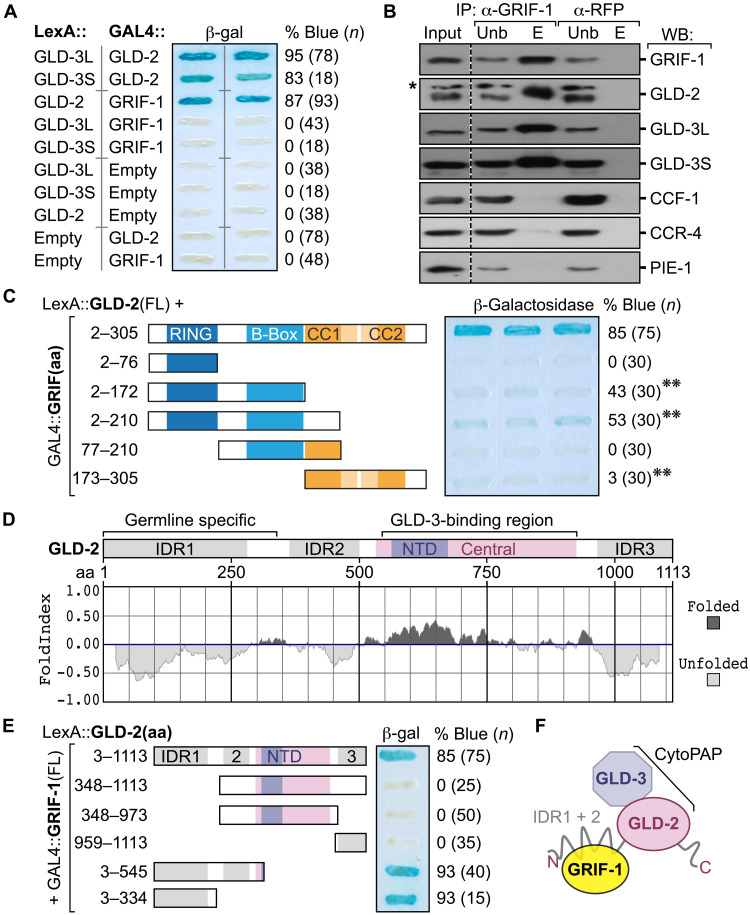
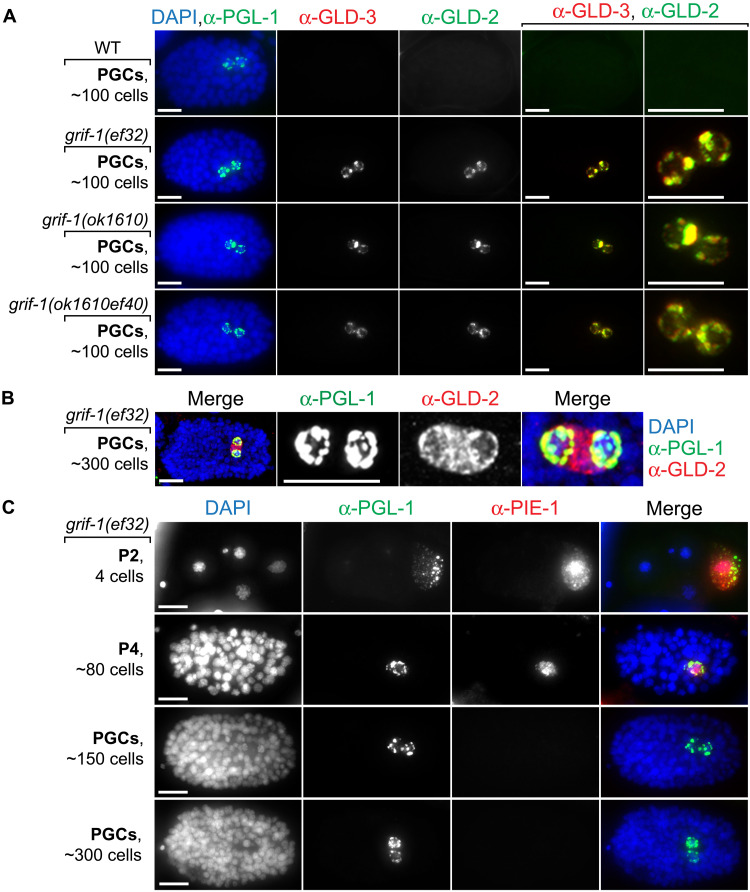
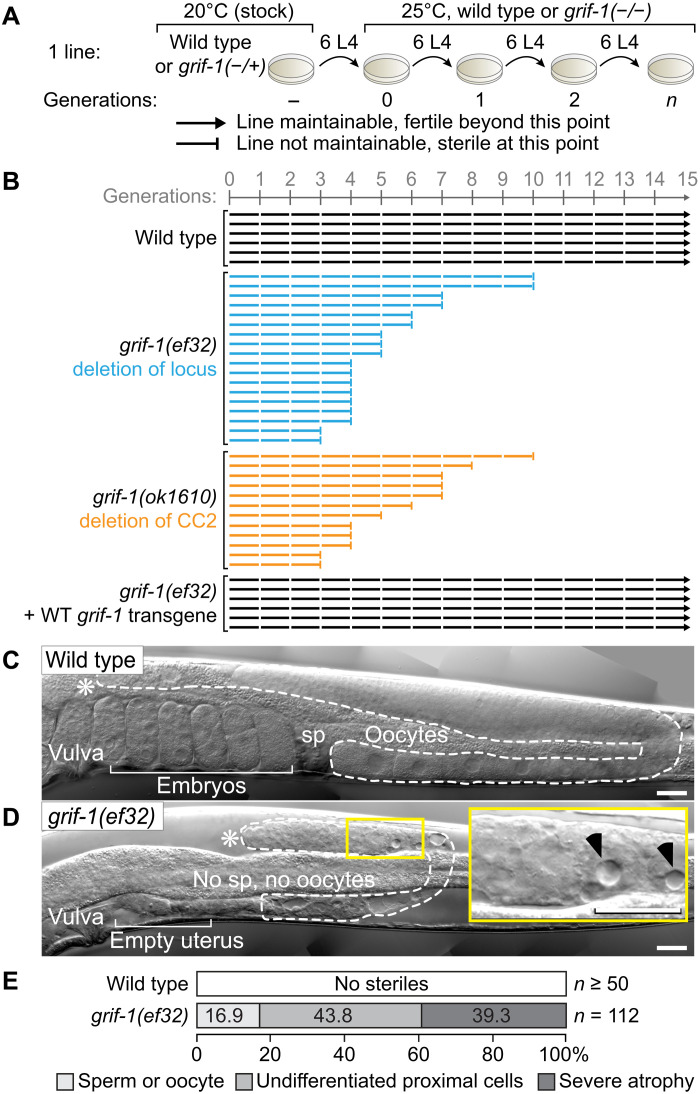
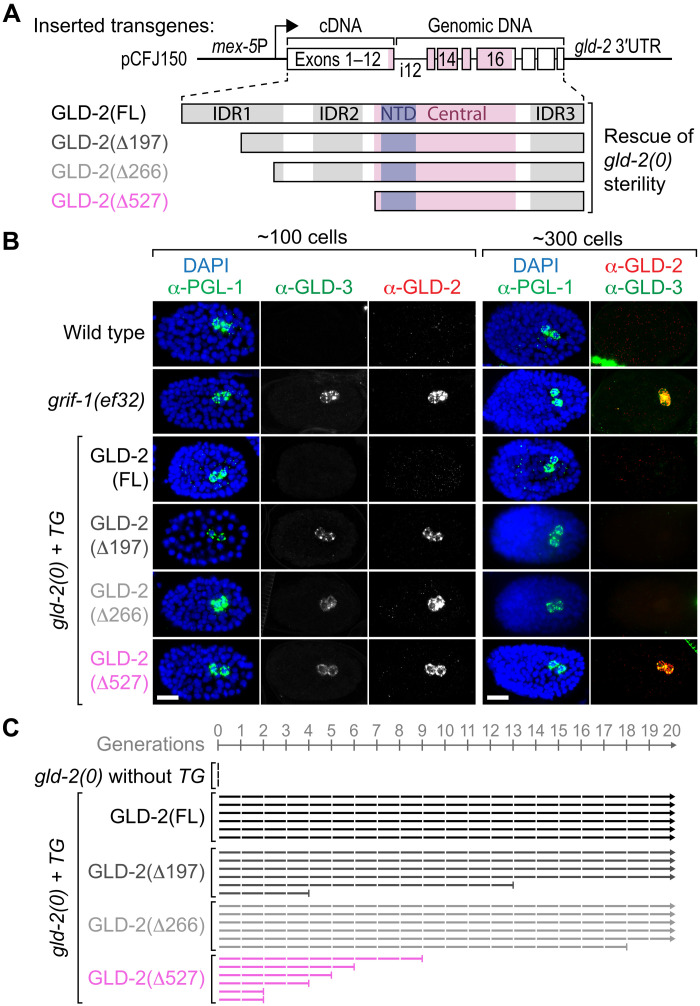
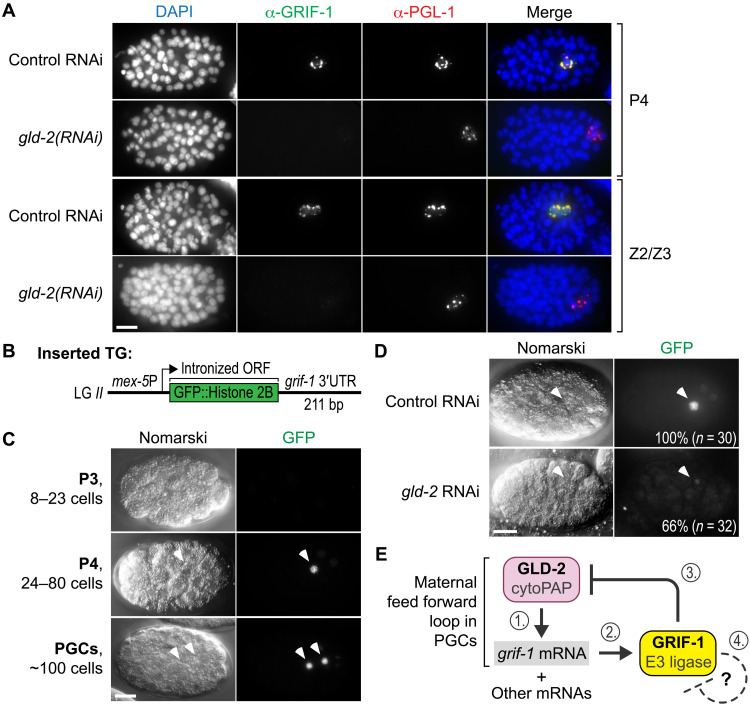
How the germ line achieves a clean transition from maternal to zygotic gene expression control is a fundamental problem in sexually reproducing organisms. Whereas several mechanisms terminate the maternal program in the soma, this combined molecular reset and handover are poorly understood for primordial germ cells (PGCs). Here, we show that GRIF-1, a TRIM32-related and presumed E3 ubiquitin ligase in Caenorhabditis elegans, eliminates the maternal cytoplasmic poly(A) polymerase (cytoPAP) complex by targeting the germline-specific intrinsically disordered region of its enzymatic subunit, GLD-2, for proteasome-mediated degradation. Interference with cytoPAP turnover in PGCs causes frequent transgenerational sterility and, eventually, germline mortality. Hence, positively acting maternal RNA regulators are cleared via the proteasome system to avoid likely interference between maternal and zygotic gene expression programs to maintain transgenerational fertility and acquire germline immortality. This strategy is likely used in all animals that preform their immortal germ line via maternally inherited germplasm determinants.
Figures
Similar articles
-
GLD-4-mediated translational activation regulates the size of the proliferative germ cell pool in the adult C. elegans germ line.PLoS Genet. 2014 Sep 25;10(9):e1004647. doi: 10.1371/journal.pgen.1004647. eCollection 2014 Sep. PLoS Genet. 2014. PMID: 25254367 Free PMC article.
-
The AAA-ATPase FIGL-1 controls mitotic progression, and its levels are regulated by the CUL-3MEL-26 E3 ligase in the C. elegans germ line.J Cell Sci. 2007 Sep 15;120(Pt 18):3179-87. doi: 10.1242/jcs.015883. J Cell Sci. 2007. PMID: 17878235
-
Proteasome regulation of the chromodomain protein MRG-1 controls the balance between proliferative fate and differentiation in the C. elegans germ line.Development. 2015 Jan 15;142(2):291-302. doi: 10.1242/dev.115147. Development. 2015. PMID: 25564623 Free PMC article.
-
Germ granules and gene regulation in the Caenorhabditis elegans germline.Genetics. 2022 Mar 3;220(3):iyab195. doi: 10.1093/genetics/iyab195. Genetics. 2022. PMID: 35239965 Free PMC article. Review.
-
Connecting the Dots: Linking Caenorhabditis elegans Small RNA Pathways and Germ Granules.Trends Cell Biol. 2021 May;31(5):387-401. doi: 10.1016/j.tcb.2020.12.012. Epub 2021 Jan 29. Trends Cell Biol. 2021. PMID: 33526340 Review.
Cited by
-
Caenorhabditis elegans germ granules accumulate hundreds of low translation mRNAs with no systematic preference for germ cell fate regulators.Development. 2024 Jul 1;151(13):dev202575. doi: 10.1242/dev.202575. Epub 2024 Jul 10. Development. 2024. PMID: 38984542 Free PMC article.
-
ZIF-1-mediated degradation of endogenous and heterologous zinc finger proteins in the C. elegans germ line.bioRxiv [Preprint]. 2023 Jul 10:2023.07.10.548405. doi: 10.1101/2023.07.10.548405. bioRxiv. 2023. Update in: Genetics. 2023 Nov 1;225(3):iyad160. doi: 10.1093/genetics/iyad160. PMID: 37502839 Free PMC article. Updated. Preprint.
-
Organogenetic transcriptomes of the Drosophila embryo at single cell resolution.Development. 2024 Jan 15;151(2):dev202097. doi: 10.1242/dev.202097. Epub 2024 Jan 18. Development. 2024. PMID: 38174902 Free PMC article.
-
ZIF-1-mediated degradation of zinc finger proteins in the Caenorhabditis elegans germ line.Genetics. 2023 Nov 1;225(3):iyad160. doi: 10.1093/genetics/iyad160. Genetics. 2023. PMID: 37647858 Free PMC article.
References
-
- Vastenhouw N. L., Cao W. X., Lipshitz H. D., The maternal-to-zygotic transition revisited. Development 146, dev161471 (2019). - PubMed
-
- Cao W. X., Kabelitz S., Gupta M., Yeung E., Lin S., Rammelt C., Ihling C., Pekovic F., Low T. C. H., Siddiqui N. U., Cheng M. H. K., Angers S., Smibert C. A., Wühr M., Wahle E., Lipshitz H. D., Precise temporal regulation of post-transcriptional repressors is required for an orderly drosophila maternal-to-zygotic transition. Cell Rep. 31, 107783 (2020). - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
