

Mechanisms of frustrated phagocytic spreading of human neutrophils on antibody-coated surfaces
- PMID: 36242516
- PMCID: PMC9748254
- DOI: 10.1016/j.bpj.2022.10.016
Mechanisms of frustrated phagocytic spreading of human neutrophils on antibody-coated surfaces
Abstract
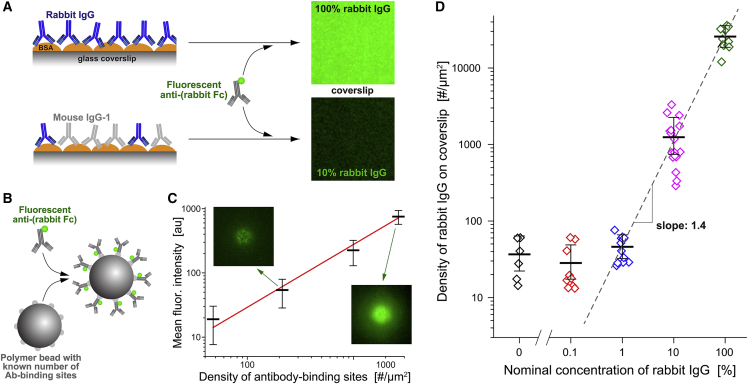
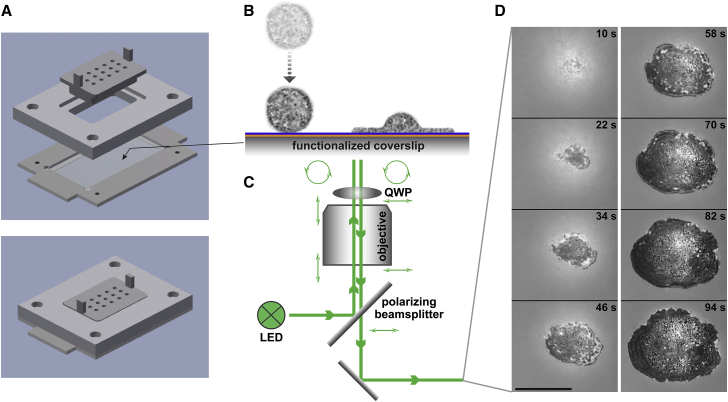
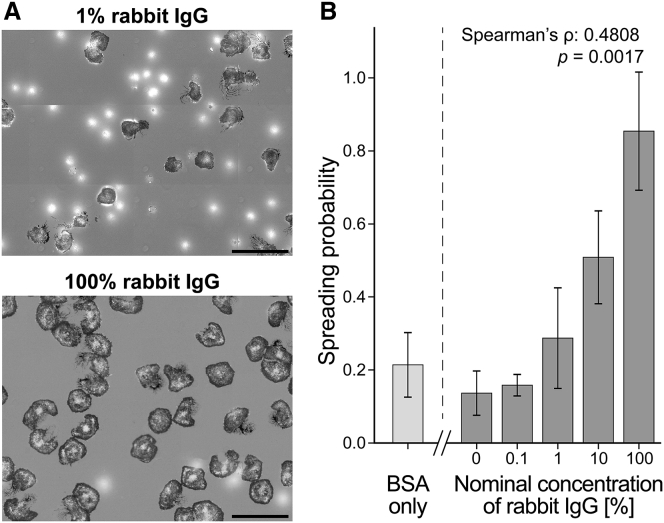
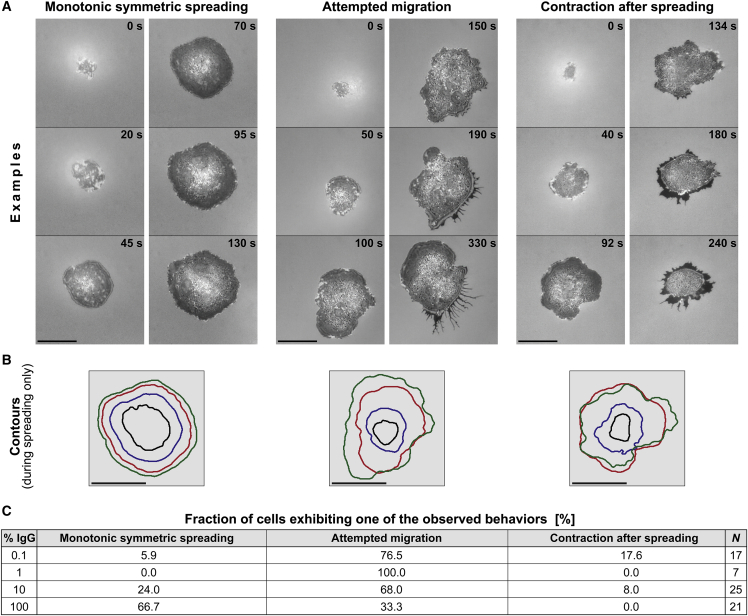
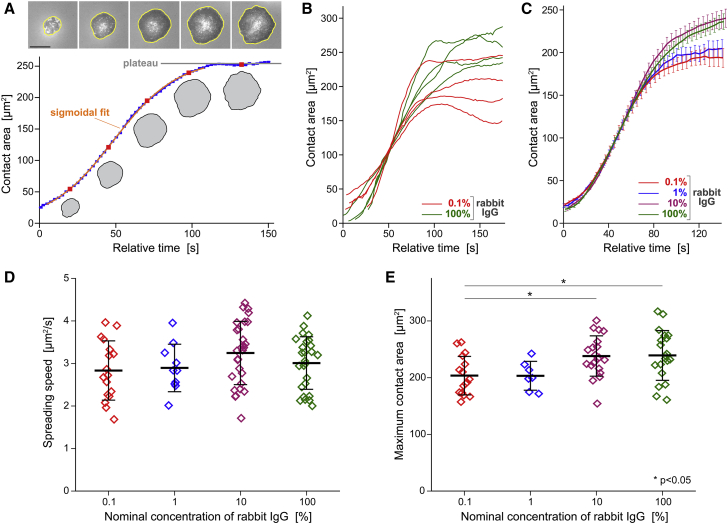
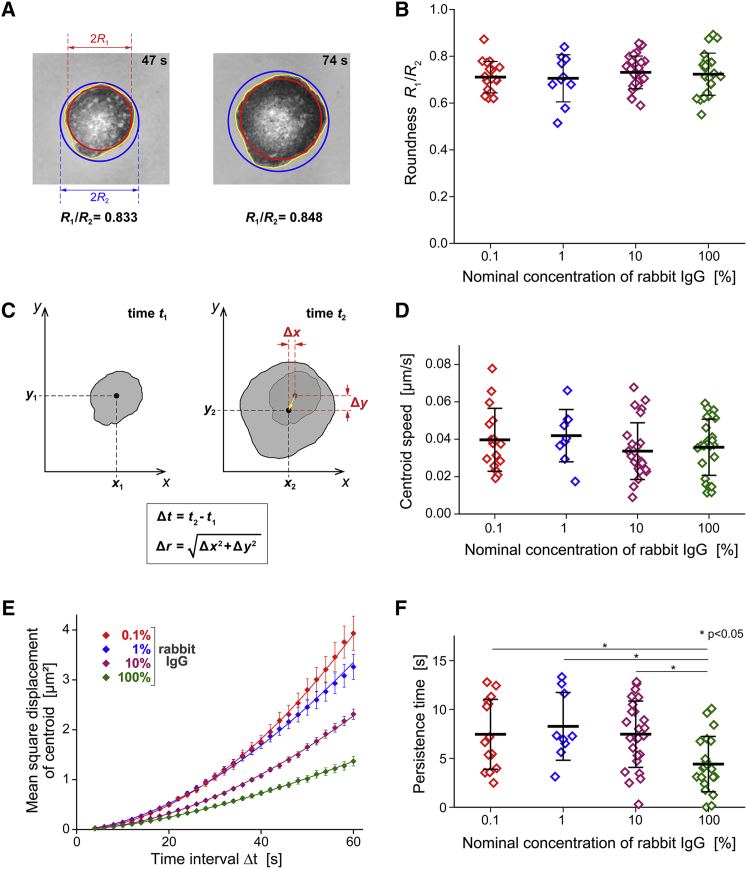
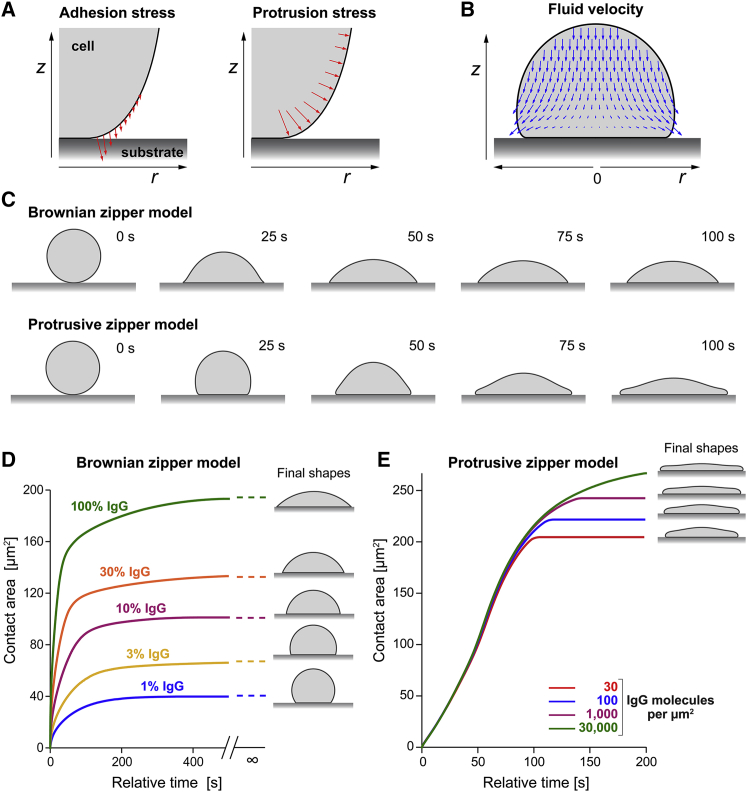
Complex motions of immune cells are an integral part of diapedesis, chemotaxis, phagocytosis, and other vital processes. To better understand how immune cells execute such motions, we present a detailed analysis of phagocytic spreading of human neutrophils on flat surfaces functionalized with different densities of immunoglobulin G (IgG) antibodies. We visualize the cell-substrate contact region at high resolution and without labels using reflection interference contrast microscopy and quantify how the area, shape, and position of the contact region evolves over time. We find that the likelihood of the cell commitment to spreading strongly depends on the surface density of IgG, but the rate at which the substrate-contact area of spreading cells increases does not. Validated by a theoretical companion study, our results resolve controversial notions about the mechanisms controlling cell spreading, establishing that active forces generated by the cytoskeleton rather than cell-substrate adhesion primarily drive cellular protrusion. Adhesion, on the other hand, aids phagocytic spreading by regulating the cell commitment to spreading, the maximum cell-substrate contact area, and the directional movement of the contact region.
Copyright © 2022 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Similar articles
-
Integrative experimental/computational approach establishes active cellular protrusion as the primary driving force of phagocytic spreading by immune cells.PLoS Comput Biol. 2022 Aug 26;18(8):e1009937. doi: 10.1371/journal.pcbi.1009937. eCollection 2022 Aug. PLoS Comput Biol. 2022. PMID: 36026476 Free PMC article.
-
Macrophages form circular zones of very close apposition to IgG-coated surfaces.Cell Motil Cytoskeleton. 1990;15(4):260-70. doi: 10.1002/cm.970150408. Cell Motil Cytoskeleton. 1990. PMID: 2337930
-
Frustrated Phagocytic Spreading of J774A-1 Macrophages Ends in Myosin II-Dependent Contraction.Biophys J. 2016 Dec 20;111(12):2698-2710. doi: 10.1016/j.bpj.2016.11.009. Biophys J. 2016. PMID: 28002746 Free PMC article.
-
A minimal cell model for lamellipodia-based cellular dynamics and migration.J Cell Sci. 2023 Jul 15;136(14):jcs260744. doi: 10.1242/jcs.260744. Epub 2023 Jul 27. J Cell Sci. 2023. PMID: 37497740 Review.
-
Blurred line between chemotactic chase and phagocytic consumption: an immunophysical single-cell perspective.J Cell Sci. 2011 Sep 15;124(Pt 18):3041-51. doi: 10.1242/jcs.086413. J Cell Sci. 2011. PMID: 21914817 Free PMC article. Review.
Cited by
-
Neutrophil Heterogeneity in Airway Inflammatory Diseases.Inflammation. 2025 Aug 19. doi: 10.1007/s10753-025-02351-z. Online ahead of print. Inflammation. 2025. PMID: 40826205 Review.
-
Integrative experimental/computational approach establishes active cellular protrusion as the primary driving force of phagocytic spreading by immune cells.PLoS Comput Biol. 2022 Aug 26;18(8):e1009937. doi: 10.1371/journal.pcbi.1009937. eCollection 2022 Aug. PLoS Comput Biol. 2022. PMID: 36026476 Free PMC article.
-
Particle-based simulations shed light on cytoskeleton-membrane dynamics in phagocytosis.Biophys J. 2024 May 7;123(9):1031-1033. doi: 10.1016/j.bpj.2024.03.032. Epub 2024 Mar 28. Biophys J. 2024. PMID: 38549374 Free PMC article. No abstract available.
-
Potent broadly neutralizing antibodies mediate efficient antibody-dependent phagocytosis of HIV-infected cells.PLoS Pathog. 2024 Oct 28;20(10):e1012665. doi: 10.1371/journal.ppat.1012665. eCollection 2024 Oct. PLoS Pathog. 2024. PMID: 39466835 Free PMC article.
-
Lacking ARHGAP25 mitigates the symptoms of autoantibody-induced arthritis in mice.Front Immunol. 2023 May 10;14:1182278. doi: 10.3389/fimmu.2023.1182278. eCollection 2023. Front Immunol. 2023. PMID: 37234175 Free PMC article.
