

Glyoxalase I activity affects Arabidopsis sensitivity to ammonium nutrition
- PMID: 36242617
- PMCID: PMC9700585
- DOI: 10.1007/s00299-022-02931-5
Glyoxalase I activity affects Arabidopsis sensitivity to ammonium nutrition
Abstract
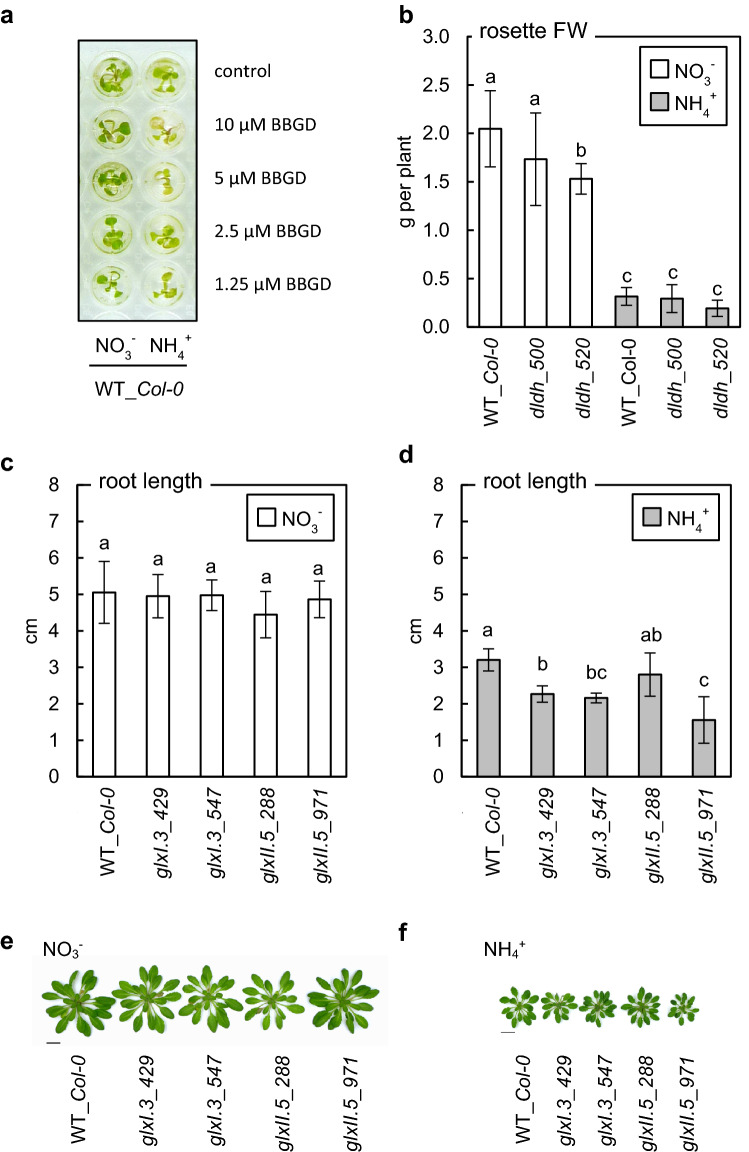
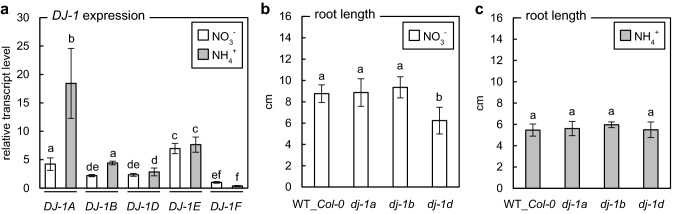
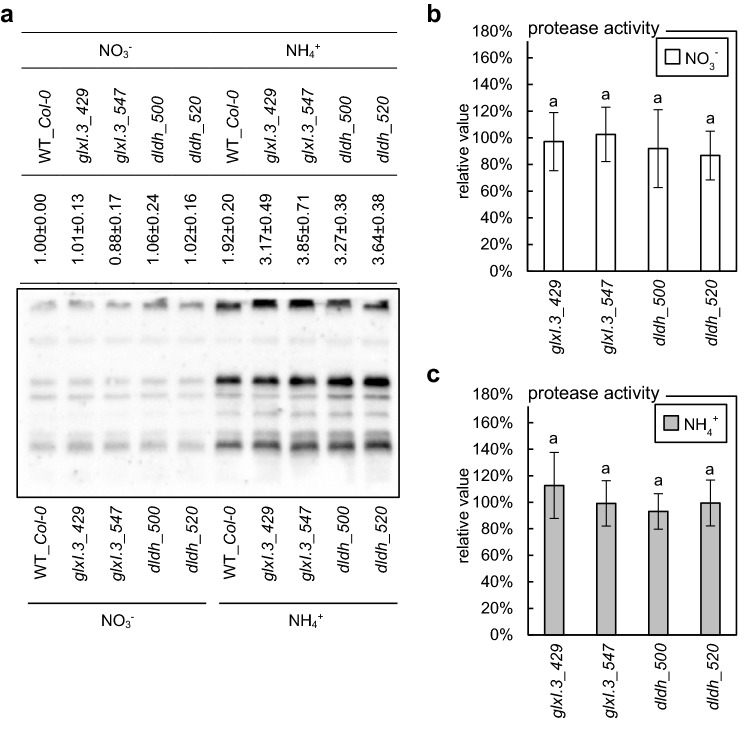
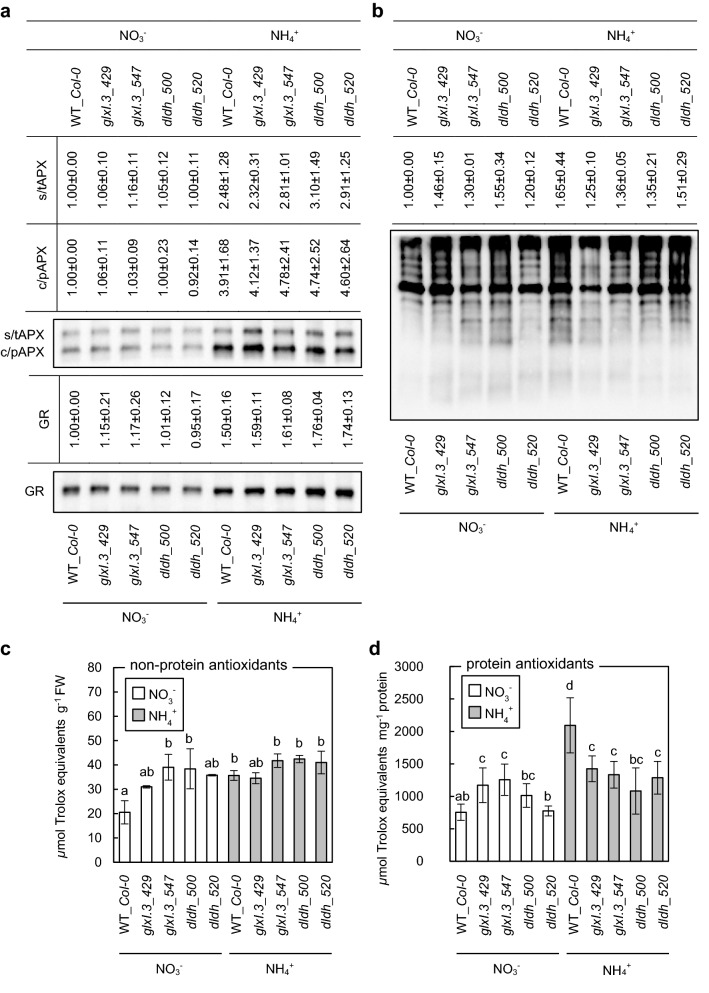
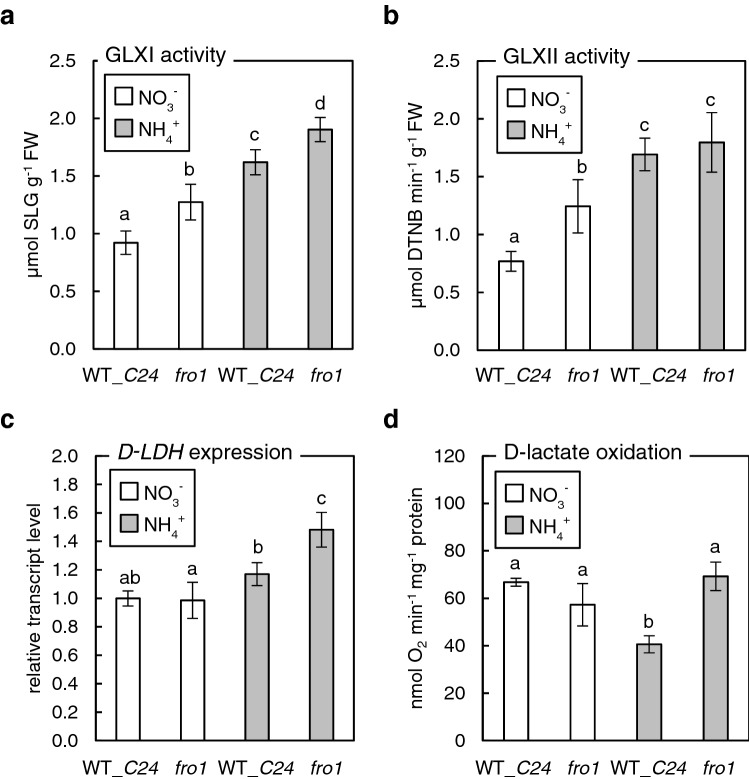
Elevated methylglyoxal levels contribute to ammonium-induced growth disorders in Arabidopsis thaliana. Methylglyoxal detoxification pathway limitation, mainly the glyoxalase I activity, leads to enhanced sensitivity of plants to ammonium nutrition. Ammonium applied to plants as the exclusive source of nitrogen often triggers multiple phenotypic effects, with severe growth inhibition being the most prominent symptom. Glycolytic flux increase, leading to overproduction of its toxic by-product methylglyoxal (MG), is one of the major metabolic consequences of long-term ammonium nutrition. This study aimed to evaluate the influence of MG metabolism on ammonium-dependent growth restriction in Arabidopsis thaliana plants. As the level of MG in plant cells is maintained by the glyoxalase (GLX) system, we analyzed MG-related metabolism in plants with a dysfunctional glyoxalase pathway. We report that MG detoxification, based on glutathione-dependent glyoxalases, is crucial for plants exposed to ammonium nutrition, and its essential role in ammonium sensitivity relays on glyoxalase I (GLXI) activity. Our results indicated that the accumulation of MG-derived advanced glycation end products significantly contributes to the incidence of ammonium toxicity symptoms. Using A. thaliana frostbite1 as a model plant that overcomes growth repression on ammonium, we have shown that its resistance to enhanced MG levels is based on increased GLXI activity and tolerance to elevated MG-derived advanced glycation end-product (MAGE) levels. Furthermore, our results show that glyoxalase pathway activity strongly affects cellular antioxidative systems. Under stress conditions, the disruption of the MG detoxification pathway limits the functioning of antioxidant defense. However, under optimal growth conditions, a defect in the MG detoxification route results in the activation of antioxidative systems.
Keywords: Ammonium nutrition; D-Lactate dehydrogenase; Dicarbonyl stress; Glyoxalase; Methylglyoxal; Mitochondrial Complex I mutant.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures





Similar articles
-
Methylglyoxal induces inhibition of growth, accumulation of anthocyanin, and activation of glyoxalase I and II in Arabidopsis thaliana.J Biochem Mol Toxicol. 2017 Jul;31(7). doi: 10.1002/jbt.21901. Epub 2017 Jan 24. J Biochem Mol Toxicol. 2017. PMID: 28117932
-
Enhanced Formation of Methylglyoxal-Derived Advanced Glycation End Products in Arabidopsis Under Ammonium Nutrition.Front Plant Sci. 2018 May 24;9:667. doi: 10.3389/fpls.2018.00667. eCollection 2018. Front Plant Sci. 2018. PMID: 29881392 Free PMC article.
-
From methylglyoxal to pyruvate: a genome-wide study for the identification of glyoxalases and D-lactate dehydrogenases in Sorghum bicolor.BMC Genomics. 2020 Feb 10;21(1):145. doi: 10.1186/s12864-020-6547-7. BMC Genomics. 2020. PMID: 32041545 Free PMC article.
-
Coordinated Actions of Glyoxalase and Antioxidant Defense Systems in Conferring Abiotic Stress Tolerance in Plants.Int J Mol Sci. 2017 Jan 20;18(1):200. doi: 10.3390/ijms18010200. Int J Mol Sci. 2017. PMID: 28117669 Free PMC article. Review.
-
Glyoxalase Goes Green: The Expanding Roles of Glyoxalase in Plants.Int J Mol Sci. 2017 Apr 24;18(4):898. doi: 10.3390/ijms18040898. Int J Mol Sci. 2017. PMID: 28441779 Free PMC article. Review.
Cited by
-
Ammonium treatment inhibits cell cycle activity and induces nuclei endopolyploidization in Arabidopsis thaliana.Planta. 2024 Mar 20;259(5):94. doi: 10.1007/s00425-024-04372-8. Planta. 2024. PMID: 38509428
-
Cobalt and Titanium Alleviate the Methylglyoxal-Induced Oxidative Stress in Pennisetum divisum Seedlings under Saline Conditions.Metabolites. 2023 Nov 19;13(11):1162. doi: 10.3390/metabo13111162. Metabolites. 2023. PMID: 37999257 Free PMC article.
-
Genome-Wide Analysis and Expression Profiling of Glyoxalase Gene Families Under Abiotic Stresses in Cucumber (Cucumis sativus L.).Int J Mol Sci. 2024 Oct 20;25(20):11294. doi: 10.3390/ijms252011294. Int J Mol Sci. 2024. PMID: 39457076 Free PMC article.
-
Genome-wide analysis and expression profiling of glyoxalase gene families in oat (Avena sativa) indicate their responses to abiotic stress during seed germination.Front Plant Sci. 2023 Jun 15;14:1215084. doi: 10.3389/fpls.2023.1215084. eCollection 2023. Front Plant Sci. 2023. PMID: 37396634 Free PMC article.
References
-
- Andreeva A, Bekkhozhin Z, Omertassova N, Baizhumanov T, Yeltay G, Akhmetali M, Toibazar D, Utepbergenov D. The apparent deglycase activity of DJ-1 results from the conversion of free methylglyoxal present in fast equilibrium with hemithioacetals and hemiaminals. J Biol Chem. 2019;294:18863–18872. doi: 10.1074/jbc.RA119.011237. - DOI - PMC - PubMed
-
- Bilova T, Lukasheva E, Brauch D, Greifenhagen U, Paudel G, Tarakhovskaya E, Frolova N, Mittasch J, Balcke GU, Tissier A, Osmolovskaya N, Vogt T, Wessjohann LA, Birkemeyer C, Milkowski C, Frolov A. A Snapshot of the plant glycated proteome. J Biol Chem. 2016;291:7621–7636. doi: 10.1074/jbc.M115.678581. - DOI - PMC - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
