

Increasing plasma L-kynurenine impairs mitochondrial oxidative phosphorylation prior to the development of atrophy in murine skeletal muscle: A pilot study
- PMID: 36246103
- PMCID: PMC9562971
- DOI: 10.3389/fphys.2022.992413
Increasing plasma L-kynurenine impairs mitochondrial oxidative phosphorylation prior to the development of atrophy in murine skeletal muscle: A pilot study
Abstract
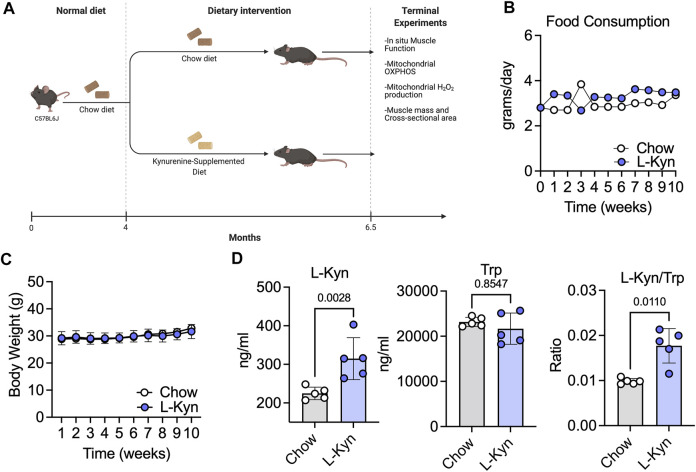
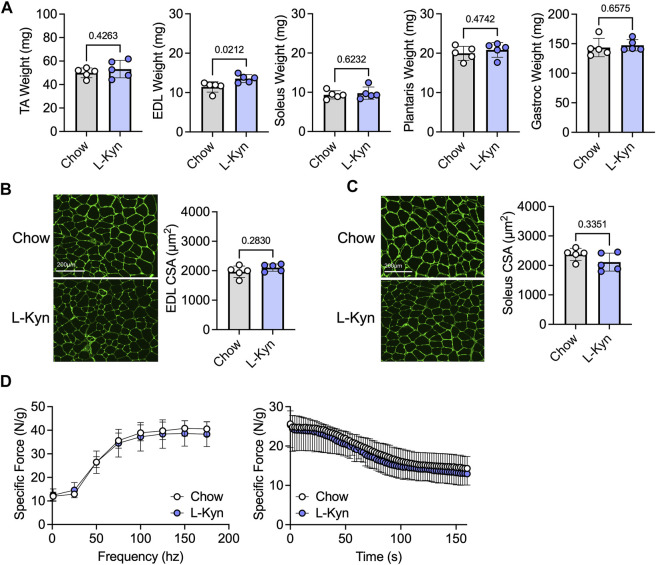
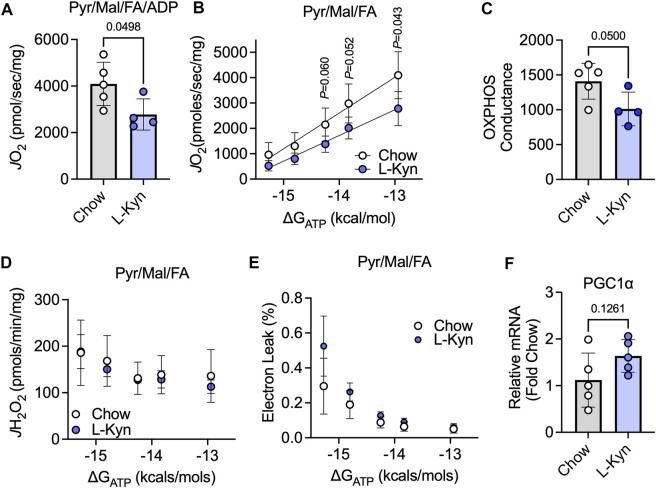
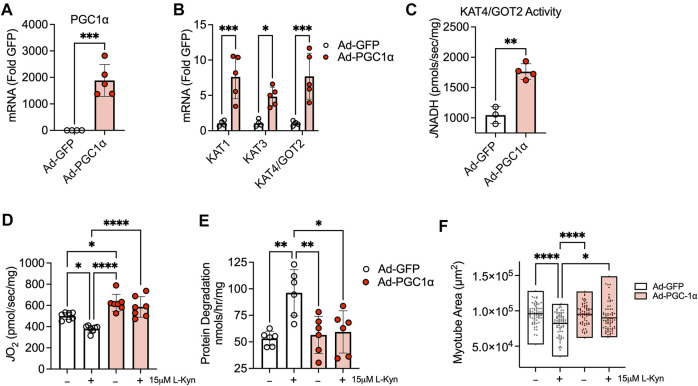
Introduction: L-Kynurenine (L-Kyn), a product of tryptophan (Trp) catabolism, has been linked with impairments in walking speed, muscle strength/size, and physical function. The purpose of this pilot study was to develop a dietary model that elevates plasma L-Kyn levels in mice and characterize its impact on muscle health and function. Methods: Four-month-old C57BL6J male mice were randomized to either a L-Kyn supplemented (150 mg/kg) or chow diet for 10 weeks. Plasma L-Kyn and Trp levels were measured via mass spectrometry. Primary outcomes included assessments of muscle weights, myofiber cross-sectional area (CSA), nerve-stimulated contractile performance, and mitochondrial oxidative phosphorylation (OXPHOS) and hydrogen peroxide (H2O2) production. Additional experiments in cultured myotubes explored the impact of enhancing L-Kyn metabolism. Results: Mice randomized to the L-Kyn diet displayed significant increases in plasma L-Kyn levels (p = 0.0028) and the L-Kyn/Trp ratio (p = 0.011) when compared to chow fed mice. Food intake and body weights were not different between groups. There were no detectable differences in muscle weights, myofiber CSA, or contractile performance. L-Kyn fed mice displayed reductions in mitochondrial OXPHOS (p = 0.05) and maximal ADP-stimulated respiration (p = 0.0498). In cultured myotubes, overexpression of peroxisome proliferator-activated receptor-gamma coactivator 1 alpha prevented atrophy and proteolysis, as well as deficits in mitochondrial respiration with L-Kyn treatment. Conclusion: Dietary feeding of L-Kyn increases plasma L-Kyn levels and the L-Kyn/Trp ratio in healthy male mice. Mitochondrial impairments in muscle were observed in mice with elevated L-Kyn without changes in muscle size or function. Enhancing L-Kyn metabolism can protect against these effects in culture myotubes.
Keywords: energetics; metabolism; mitochondria; physical function; weakness.
Copyright © 2022 Palzkill, Thome, Murillo, Khattri and Ryan.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
Similar articles
-
The aromatic amino acid tryptophan stimulates skeletal muscle IGF1/p70s6k/mTor signaling in vivo and the expression of myogenic genes in vitro.Nutrition. 2015 Jul-Aug;31(7-8):1018-24. doi: 10.1016/j.nut.2015.02.011. Epub 2015 Mar 17. Nutrition. 2015. PMID: 26059377 Free PMC article.
-
The Power Struggle: Kynurenine Pathway Enzyme Knockouts and Brain Mitochondrial Respiration.J Neurochem. 2025 May;169(5):e70075. doi: 10.1111/jnc.70075. J Neurochem. 2025. PMID: 40317489 Free PMC article.
-
Kynurenine metabolism is altered in mdx mice: a potential muscle to brain connection.Exp Physiol. 2022 Sep;107(9):1029-1036. doi: 10.1113/EP090381. Epub 2022 Aug 15. Exp Physiol. 2022. PMID: 35912981
-
Mitochondria, Oxidative Stress and the Kynurenine System, with a Focus on Ageing and Neuroprotection.Molecules. 2018 Jan 17;23(1):191. doi: 10.3390/molecules23010191. Molecules. 2018. PMID: 29342113 Free PMC article. Review.
-
The tryptophan catabolite or kynurenine pathway in schizophrenia: meta-analysis reveals dissociations between central, serum, and plasma compartments.Mol Psychiatry. 2022 Sep;27(9):3679-3691. doi: 10.1038/s41380-022-01552-4. Epub 2022 Apr 14. Mol Psychiatry. 2022. PMID: 35422466
Cited by
-
Review of Exercise Interventions to Improve Clinical Outcomes in Nondialysis CKD.Kidney Int Rep. 2024 Aug 2;9(11):3097-3115. doi: 10.1016/j.ekir.2024.07.032. eCollection 2024 Nov. Kidney Int Rep. 2024. PMID: 39534200 Free PMC article. Review.
-
Glucocorticoid-Mediated Skeletal Muscle Atrophy: Molecular Mechanisms and Potential Therapeutic Targets.Int J Mol Sci. 2025 Aug 6;26(15):7616. doi: 10.3390/ijms26157616. Int J Mol Sci. 2025. PMID: 40806744 Free PMC article. Review.
-
Redefining Roles: A Paradigm Shift in Tryptophan-Kynurenine Metabolism for Innovative Clinical Applications.Int J Mol Sci. 2024 Nov 27;25(23):12767. doi: 10.3390/ijms252312767. Int J Mol Sci. 2024. PMID: 39684480 Free PMC article. Review.
-
Gut microbiota and the tryptophan-kynurenine pathway in anxiety: new insights and treatment strategies.J Neural Transm (Vienna). 2025 Jul;132(7):943-977. doi: 10.1007/s00702-025-02938-8. Epub 2025 May 14. J Neural Transm (Vienna). 2025. PMID: 40369368 Review.
-
The Biology and Biochemistry of Kynurenic Acid, a Potential Nutraceutical with Multiple Biological Effects.Int J Mol Sci. 2024 Aug 21;25(16):9082. doi: 10.3390/ijms25169082. Int J Mol Sci. 2024. PMID: 39201768 Free PMC article. Review.
References
Grants and funding
LinkOut - more resources
Full Text Sources