

Isoprenoid biosynthesis regulation in poplars by methylerythritol phosphate and mevalonic acid pathways
- PMID: 36247639
- PMCID: PMC9562105
- DOI: 10.3389/fpls.2022.968780
Isoprenoid biosynthesis regulation in poplars by methylerythritol phosphate and mevalonic acid pathways
Abstract
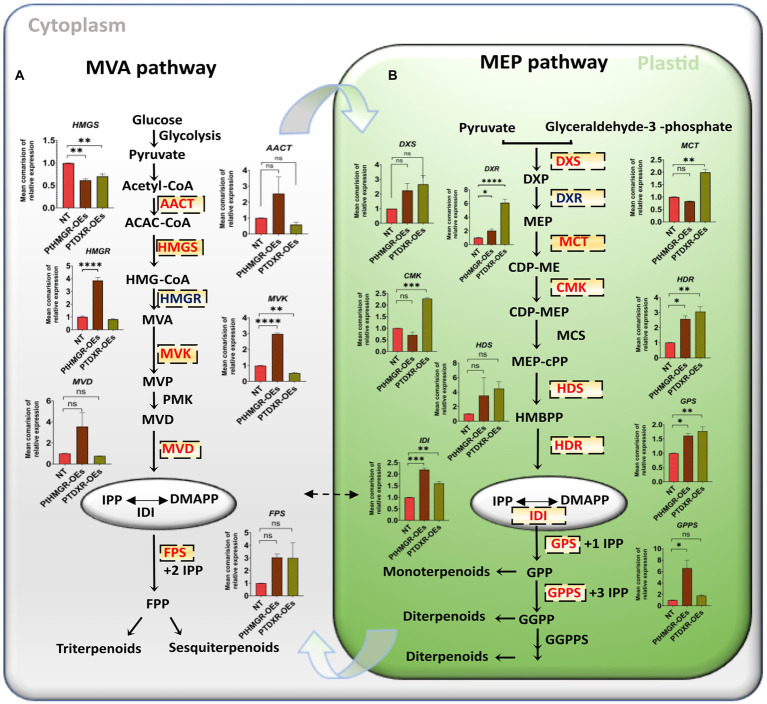
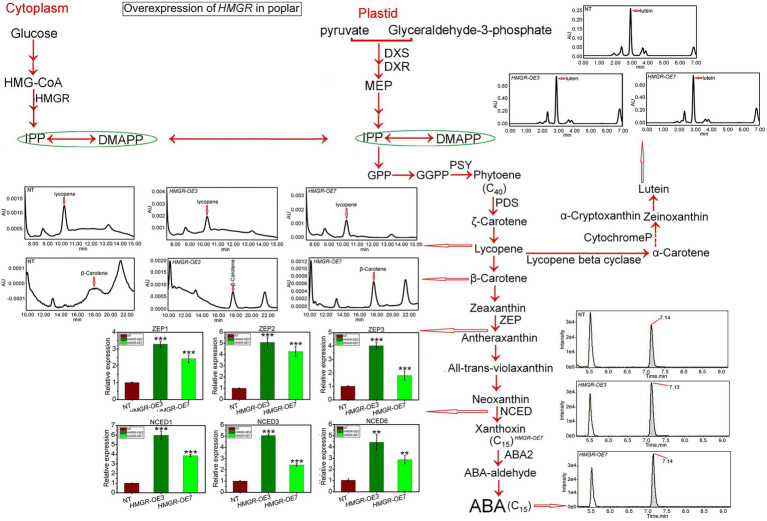
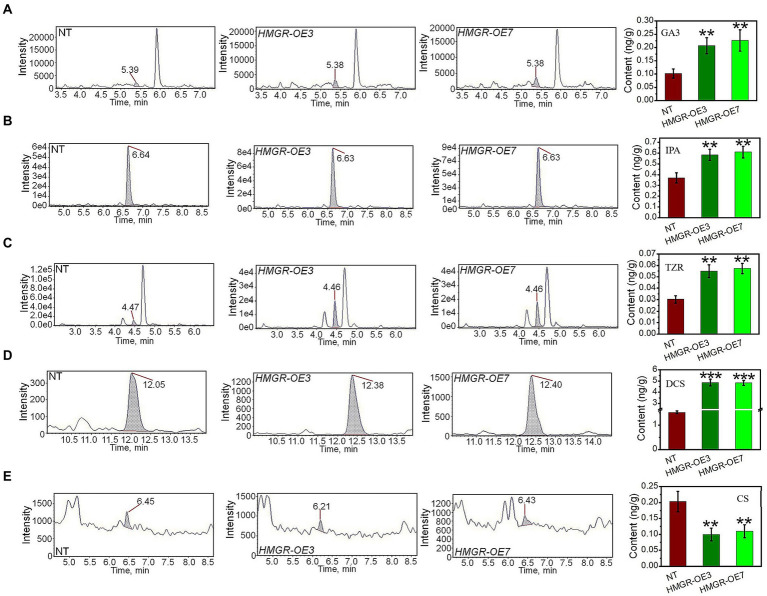
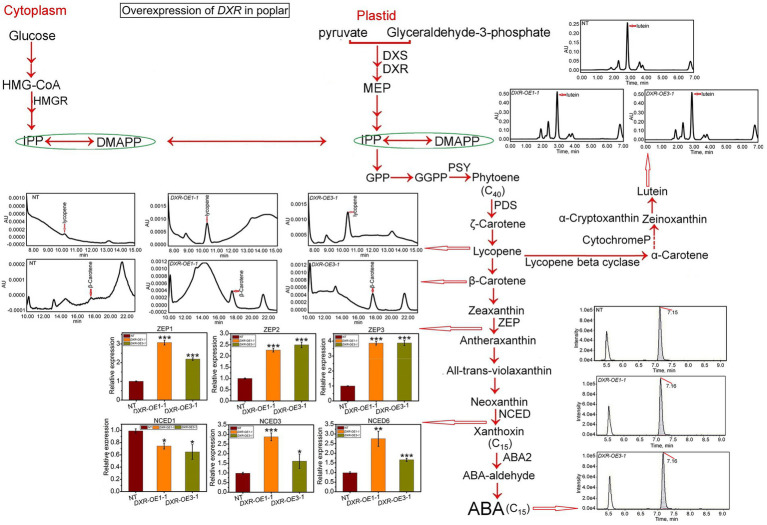
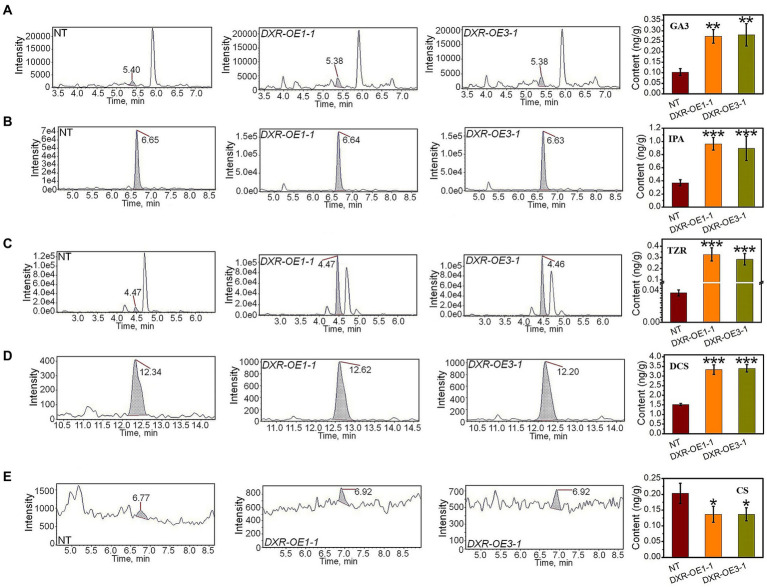
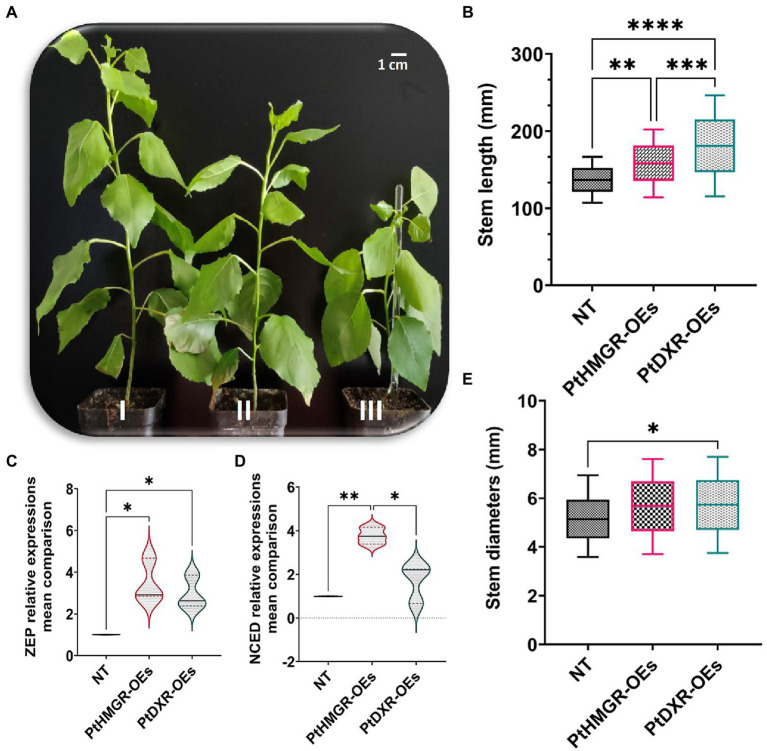
It is critical to develop plant isoprenoid production when dealing with human-demanded industries such as flavoring, aroma, pigment, pharmaceuticals, and biomass used for biofuels. The methylerythritol phosphate (MEP) and mevalonic acid (MVA) plant pathways contribute to the dynamic production of isoprenoid compounds. Still, the cross-talk between MVA and MEP in isoprenoid biosynthesis is not quite recognized. Regarding the rate-limiting steps in the MEP pathway through catalyzing 1-deoxy-D-xylulose5-phosphate synthase and 1-deoxy-D-xylulose5-phosphate reductoisomerase (DXR) and also the rate-limiting step in the MVA pathway through catalyzing 3-hydroxy-3-methylglutaryl-CoA reductase (HMGR), the characterization and function of HMGR from Populus trichocarpa (PtHMGR) were analyzed. The results indicated that PtHMGR overexpressors (OEs) displayed various MEP and MVA-related gene expressions compared to NT poplars. The overexpression of PtDXR upregulated MEP-related genes and downregulated MVA-related genes. The overexpression of PtDXR and PtHMGR affected the isoprenoid production involved in both MVA and MEP pathways. Here, results illustrated that the PtHMGR and PtDXR play significant roles in regulating MEP and MVA-related genes and derived isoprenoids. This study clarifies cross-talk between MVA and MEP pathways. It demonstrates the key functions of HMGR and DXR in this cross-talk, which significantly contribute to regulate isoprenoid biosynthesis in poplars.
Keywords: DXR; HMGR; isoprenoid biosynthesis; methylerythritol phosphate pathway; mevalonic acid pathway; poplar.
Copyright © 2022 Movahedi, Wei, Pucker, Ghaderi-Zefrehei, Rasouli, Kiani-Pouya, Jiang, Zhuge, Yang and Zhou.
Conflict of interest statement
The authors declare that this research was conducted without any commercial or financial relationships construed as a potential conflict of interest.
Figures
Similar articles
-
Characterization and Function of 3-Hydroxy-3-Methylglutaryl-CoA Reductase in Populus trichocarpa: Overexpression of PtHMGR Enhances Terpenoids in Transgenic Poplar.Front Plant Sci. 2019 Nov 15;10:1476. doi: 10.3389/fpls.2019.01476. eCollection 2019. Front Plant Sci. 2019. PMID: 31803212 Free PMC article.
-
Both methylerythritol phosphate and mevalonate pathways contribute to biosynthesis of each of the major isoprenoid classes in young cotton seedlings.Phytochemistry. 2014 Feb;98:110-9. doi: 10.1016/j.phytochem.2013.11.010. Epub 2013 Dec 17. Phytochemistry. 2014. PMID: 24359633
-
Different roles of the mevalonate and methylerythritol phosphate pathways in cell growth and tanshinone production of Salvia miltiorrhiza hairy roots.PLoS One. 2012;7(11):e46797. doi: 10.1371/journal.pone.0046797. Epub 2012 Nov 29. PLoS One. 2012. PMID: 23209548 Free PMC article.
-
Methylerythritol phosphate pathway of isoprenoid biosynthesis.Annu Rev Biochem. 2013;82:497-530. doi: 10.1146/annurev-biochem-052010-100934. Annu Rev Biochem. 2013. PMID: 23746261 Free PMC article. Review.
-
An update on the function and regulation of methylerythritol phosphate and mevalonate pathways and their evolutionary dynamics.J Integr Plant Biol. 2021 Jul;63(7):1211-1226. doi: 10.1111/jipb.13076. Epub 2021 Apr 14. J Integr Plant Biol. 2021. PMID: 33538411 Review.
Cited by
-
De Novo Transcriptome Profiling for the Generation and Validation of Microsatellite Markers, Transcription Factors, and Database Development for Andrographis paniculata.Int J Mol Sci. 2023 May 24;24(11):9212. doi: 10.3390/ijms24119212. Int J Mol Sci. 2023. PMID: 37298166 Free PMC article.
-
Integrated Analysis of Terpenoid Profiles and Full-Length Transcriptome Reveals the Central Pathways of Sesquiterpene Biosynthesis in Atractylodes chinensis (DC.) Koidz.Int J Mol Sci. 2025 Jan 26;26(3):1074. doi: 10.3390/ijms26031074. Int J Mol Sci. 2025. PMID: 39940836 Free PMC article.
-
Metabolite Profiling to Evaluate Metabolic Changes in Genetically Modified Protopanaxadiol-Enriched Rice.Plants (Basel). 2023 Feb 8;12(4):758. doi: 10.3390/plants12040758. Plants (Basel). 2023. PMID: 36840106 Free PMC article.
-
An efficient harvesting strategy for agarwood based on the correlation analysis of resin formation and leaves dynamic changes induced by integrated induction method.PLoS One. 2025 Jul 10;20(7):e0327516. doi: 10.1371/journal.pone.0327516. eCollection 2025. PLoS One. 2025. PMID: 40638569 Free PMC article.
-
Biosynthesis and the Transcriptional Regulation of Terpenoids in Tea Plants (Camellia sinensis).Int J Mol Sci. 2023 Apr 8;24(8):6937. doi: 10.3390/ijms24086937. Int J Mol Sci. 2023. PMID: 37108101 Free PMC article. Review.
References
LinkOut - more resources
Full Text Sources
Research Materials