

Transcriptional states of CAR-T infusion relate to neurotoxicity - lessons from high-resolution single-cell SOM expression portraying
- PMID: 36248848
- PMCID: PMC9558919
- DOI: 10.3389/fimmu.2022.994885
Transcriptional states of CAR-T infusion relate to neurotoxicity - lessons from high-resolution single-cell SOM expression portraying
Abstract
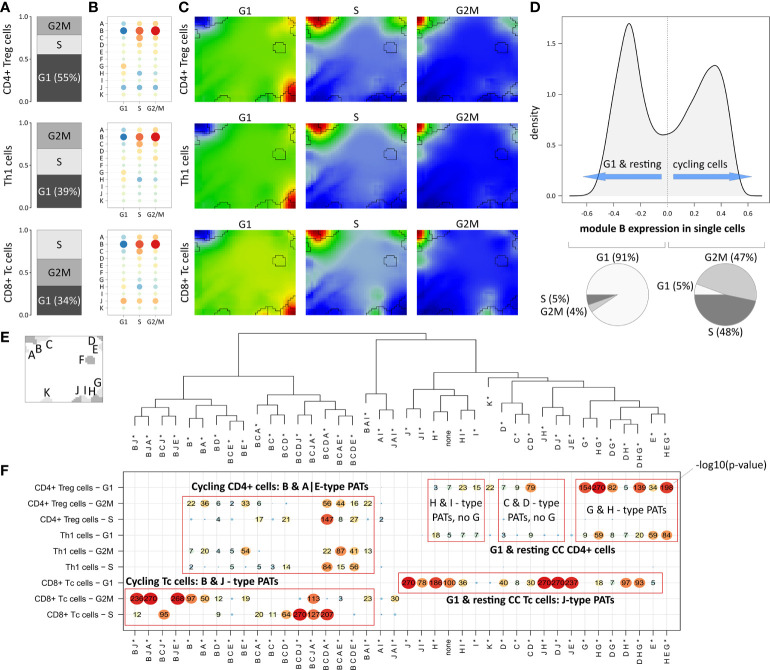
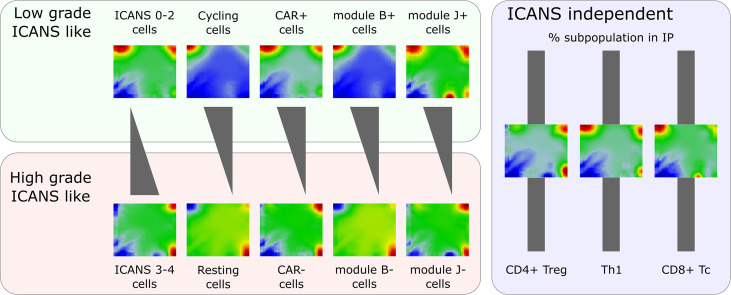
Anti-CD19 CAR-T cell immunotherapy is a hopeful treatment option for patients with B cell lymphomas, however it copes with partly severe adverse effects like neurotoxicity. Single-cell resolved molecular data sets in combination with clinical parametrization allow for comprehensive characterization of cellular subpopulations, their transcriptomic states, and their relation to the adverse effects. We here present a re-analysis of single-cell RNA sequencing data of 24 patients comprising more than 130,000 cells with focus on cellular states and their association to immune cell related neurotoxicity. For this, we developed a single-cell data portraying workflow to disentangle the transcriptional state space with single-cell resolution and its analysis in terms of modularly-composed cellular programs. We demonstrated capabilities of single-cell data portraying to disentangle transcriptional states using intuitive visualization, functional mining, molecular cell stratification, and variability analyses. Our analysis revealed that the T cell composition of the patient's infusion product as well as the spectrum of their transcriptional states of cells derived from patients with low ICANS grade do not markedly differ from those of cells from high ICANS patients, while the relative abundancies, particularly that of cycling cells, of LAG3-mediated exhaustion and of CAR positive cells, vary. Our study provides molecular details of the transcriptomic landscape with possible impact to overcome neurotoxicity.
Keywords: CAR-T cell immunotherapy; bioinformatics workflow; data portraying; single-cell transcriptomics; transcriptional states.
Copyright © 2022 Loeffler-Wirth, Rade, Arakelyan, Kreuz, Loeffler, Koehl, Reiche and Binder.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures







Similar articles
-
Single-cell multiomics dissection of basal and antigen-specific activation states of CD19-targeted CAR T cells.J Immunother Cancer. 2021 May;9(5):e002328. doi: 10.1136/jitc-2020-002328. J Immunother Cancer. 2021. PMID: 34006631 Free PMC article.
-
Imaging-based Toxicity and Response Pattern Assessment Following CAR T-Cell Therapy.Radiology. 2022 Feb;302(2):438-445. doi: 10.1148/radiol.2021210760. Epub 2021 Nov 9. Radiology. 2022. PMID: 34751616
-
Neurotoxicity-associated sinus bradycardia after chimeric antigen receptor T-cell therapy.Hematol Oncol. 2022 Aug;40(3):482-487. doi: 10.1002/hon.2976. Epub 2022 Feb 16. Hematol Oncol. 2022. PMID: 35139240
-
Immune effector cell associated neurotoxicity syndrome in chimeric antigen receptor-T cell therapy.Front Immunol. 2022 Aug 23;13:879608. doi: 10.3389/fimmu.2022.879608. eCollection 2022. Front Immunol. 2022. PMID: 36081506 Free PMC article. Review.
-
Chimeric Antigen Receptor T-Cell Therapies for Aggressive B-Cell Lymphomas: Current and Future State of the Art.Am Soc Clin Oncol Educ Book. 2019 Jan;39:446-453. doi: 10.1200/EDBK_238693. Epub 2019 May 17. Am Soc Clin Oncol Educ Book. 2019. PMID: 31099671 Review.
Cited by
-
New regulatory thinking is needed for AI-based personalised drug and cell therapies in precision oncology.NPJ Precis Oncol. 2024 Jan 30;8(1):23. doi: 10.1038/s41698-024-00517-w. NPJ Precis Oncol. 2024. PMID: 38291217 Free PMC article. Review.
-
Insights gained from single-cell analysis of chimeric antigen receptor T-cell immunotherapy in cancer.Mil Med Res. 2023 Nov 8;10(1):52. doi: 10.1186/s40779-023-00486-4. Mil Med Res. 2023. PMID: 37941075 Free PMC article. Review.
-
A Spatial Transcriptomics Browser for Discovering Gene Expression Landscapes across Microscopic Tissue Sections.Curr Issues Mol Biol. 2024 May 13;46(5):4701-4720. doi: 10.3390/cimb46050284. Curr Issues Mol Biol. 2024. PMID: 38785552 Free PMC article.
-
Transcriptomic Approaches in Studies on and Applications of Chimeric Antigen Receptor T Cells.Biomedicines. 2023 Apr 6;11(4):1107. doi: 10.3390/biomedicines11041107. Biomedicines. 2023. PMID: 37189725 Free PMC article. Review.
-
Harnessing the Transcriptional Signatures of CAR-T-Cells and Leukemia/Lymphoma Using Single-Cell Sequencing Technologies.Int J Mol Sci. 2024 Feb 19;25(4):2416. doi: 10.3390/ijms25042416. Int J Mol Sci. 2024. PMID: 38397092 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources