

Conservation of transcriptional regulation by BRCA1 and BARD1 in Caenorhabditis elegans
- PMID: 36250637
- PMCID: PMC10018340
- DOI: 10.1093/nar/gkac877
Conservation of transcriptional regulation by BRCA1 and BARD1 in Caenorhabditis elegans
Abstract
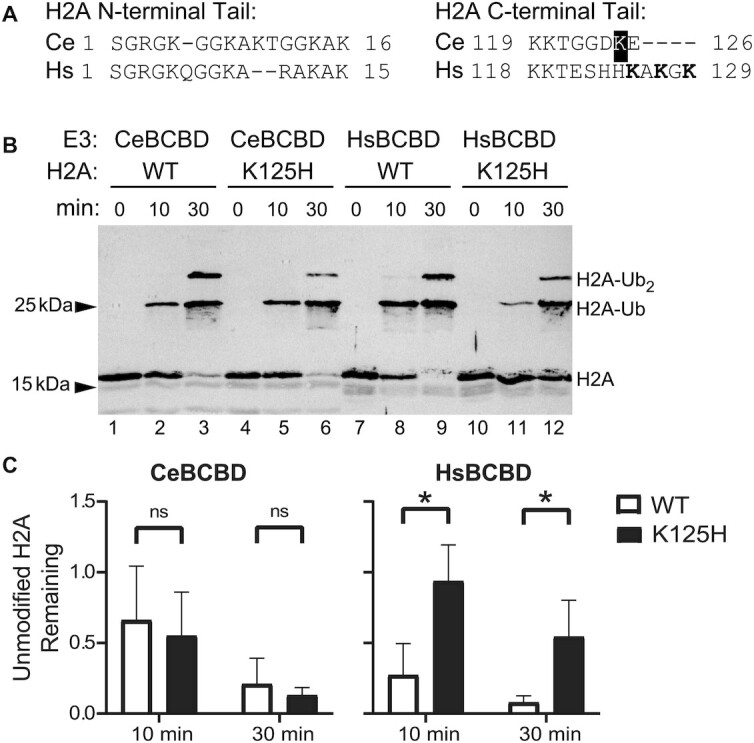
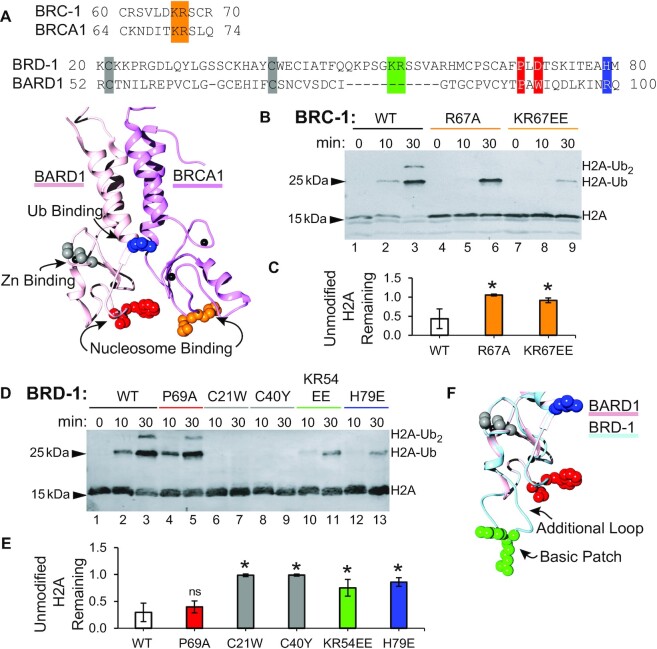
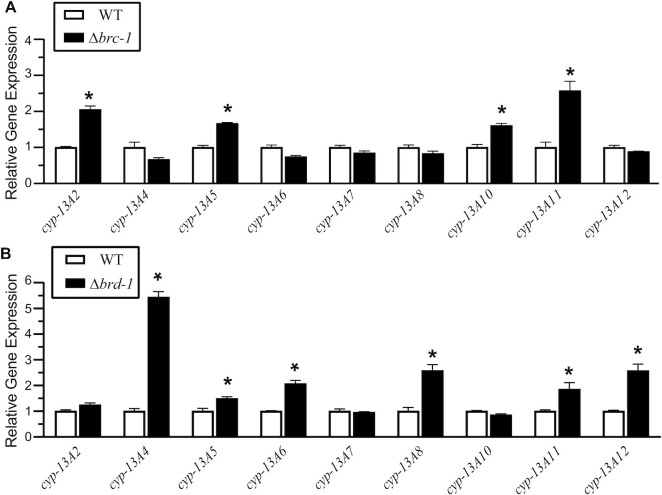
The tumor-suppressor proteins BRCA1 and BARD1 function as an E3 ubiquitin ligase to facilitate transcriptional repression and DNA damage repair. This is mediated in-part through its ability to mono-ubiquitylate histone H2A in nucleosomes. Studies in Caenorhabditis elegans have been used to elucidate numerous functions of BRCA1 and BARD1; however, it has not been established that the C. elegans orthologs, BRC-1 and BRD-1, retain all the functions of their human counterparts. Here we explore the conservation of enzymatic activity toward nucleosomes which leads to repression of estrogen-metabolizing cytochrome P450 (cyp) genes in humans. Biochemical assays establish that BRC-1 and BRD-1 contribute to ubiquitylation of histone H2A in the nucleosome. Mutational analysis shows that while BRC-1 likely binds the nucleosome using a conserved interface, BRD-1 and BARD1 have evolved different modes of binding, resulting in a difference in the placement of ubiquitin on H2A. Gene expression analysis reveals that in spite of this difference, BRC-1 and BRD-1 also contribute to cyp gene repression in C. elegans. Establishing conservation of these functions in C. elegans allows for use of this powerful model organism to address remaining questions regarding regulation of gene expression by BRCA1 and BARD1.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Snouwaert J.N., Gowen L.C., Latour A.M., Mohn A.R., Xiao A., DiBiase L., Koller B.H.. BRCA1 deficient embryonic stem cells display a decreased homologous recombination frequency and an increased frequency of non-homologous recombination that is corrected by expression of a brca1 transgene. Oncogene. 1999; 18:7900–7907. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
