

A blast fungus zinc-finger fold effector binds to a hydrophobic pocket in host Exo70 proteins to modulate immune recognition in rice
- PMID: 36252011
- PMCID: PMC9618136
- DOI: 10.1073/pnas.2210559119
A blast fungus zinc-finger fold effector binds to a hydrophobic pocket in host Exo70 proteins to modulate immune recognition in rice
Abstract
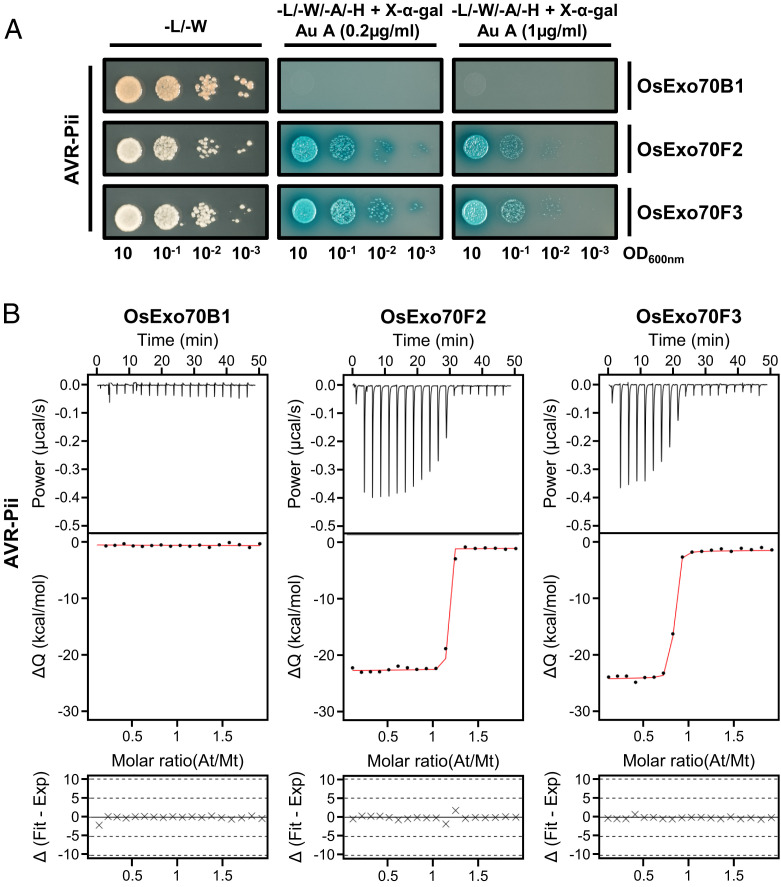
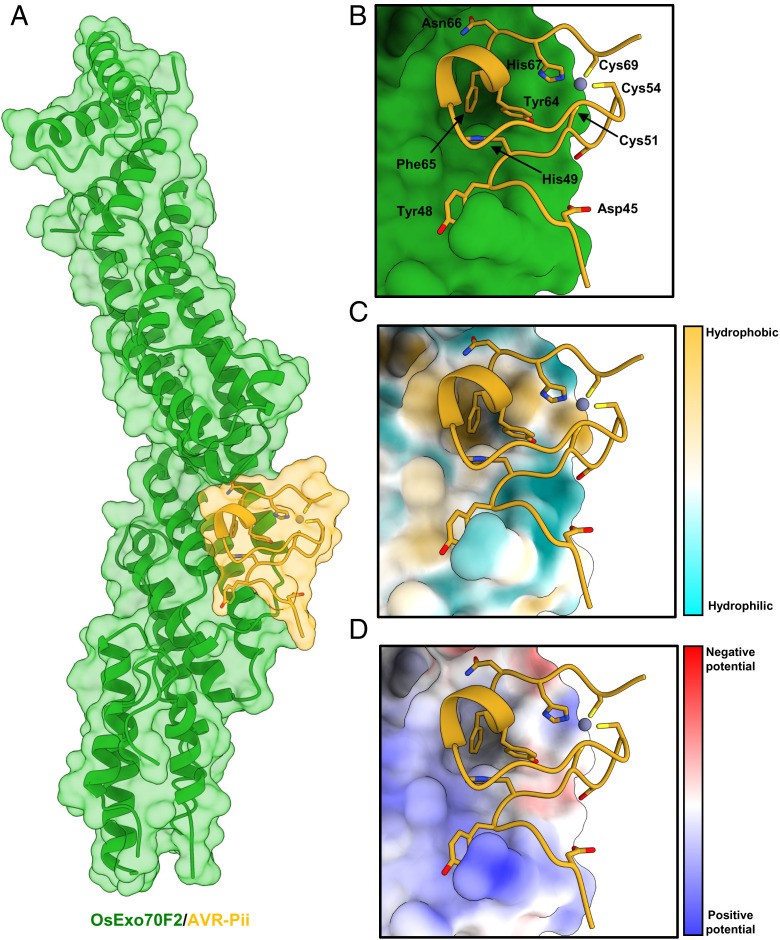
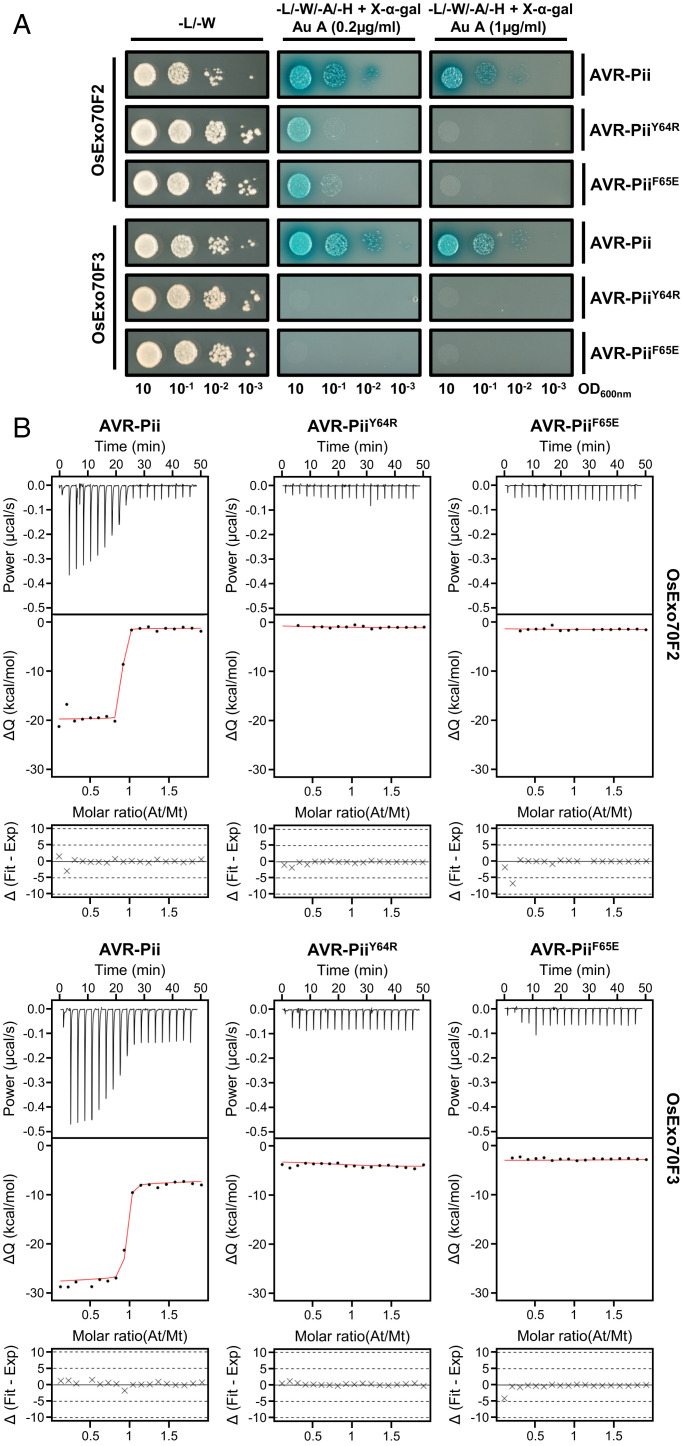
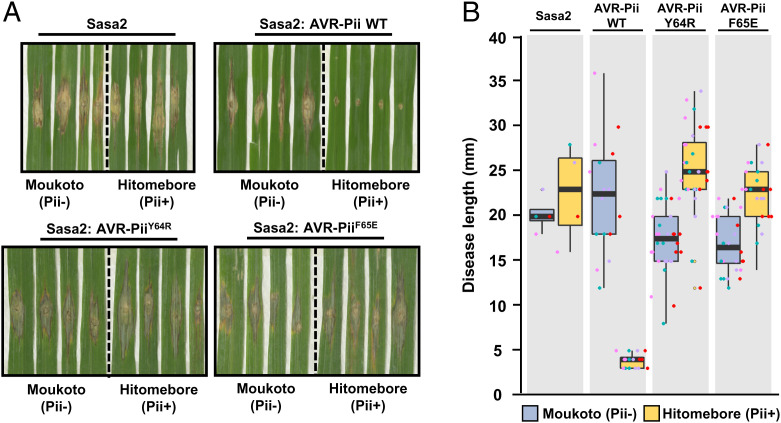
Exocytosis plays an important role in plant-microbe interactions, in both pathogenesis and symbiosis. Exo70 proteins are integral components of the exocyst, an octameric complex that mediates tethering of vesicles to membranes in eukaryotes. Although plant Exo70s are known to be targeted by pathogen effectors, the underpinning molecular mechanisms and the impact of this interaction on infection are poorly understood. Here, we show the molecular basis of the association between the effector AVR-Pii of the blast fungus Maganaporthe oryzae and rice Exo70 alleles OsExo70F2 and OsExo70F3, which is sensed by the immune receptor pair Pii via an integrated RIN4/NOI domain. The crystal structure of AVR-Pii in complex with OsExo70F2 reveals that the effector binds to a conserved hydrophobic pocket in Exo70, defining an effector/target binding interface. Structure-guided and random mutagenesis validates the importance of AVR-Pii residues at the Exo70 binding interface to sustain protein association and disease resistance in rice when challenged with fungal strains expressing effector mutants. Furthermore, the structure of AVR-Pii defines a zinc-finger effector fold (ZiF) distinct from the MAX (Magnaporthe Avrs and ToxB-like) fold previously described for a majority of characterized M. oryzae effectors. Our data suggest that blast fungus ZiF effectors bind a conserved Exo70 interface to manipulate plant exocytosis and that these effectors are also baited by plant immune receptors, pointing to new opportunities for engineering disease resistance.
Keywords: Exocyst; NLR; effector; plant immunity.
Conflict of interest statement
The authors declare no competing interest.
Figures
Similar articles
-
Zinc-finger (ZiF) fold secreted effectors form a functionally diverse family across lineages of the blast fungus Magnaporthe oryzae.PLoS Pathog. 2024 Jun 17;20(6):e1012277. doi: 10.1371/journal.ppat.1012277. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38885263 Free PMC article.
-
Specific recognition of two MAX effectors by integrated HMA domains in plant immune receptors involves distinct binding surfaces.Proc Natl Acad Sci U S A. 2018 Nov 6;115(45):11637-11642. doi: 10.1073/pnas.1810705115. Epub 2018 Oct 24. Proc Natl Acad Sci U S A. 2018. PMID: 30355769 Free PMC article.
-
Cross-reactivity of a rice NLR immune receptor to distinct effectors from the rice blast pathogen Magnaporthe oryzae provides partial disease resistance.J Biol Chem. 2019 Aug 30;294(35):13006-13016. doi: 10.1074/jbc.RA119.007730. Epub 2019 Jul 11. J Biol Chem. 2019. PMID: 31296569 Free PMC article.
-
Effector-triggered susceptibility by the rice blast fungus Magnaporthe oryzae.New Phytol. 2024 Feb;241(3):1007-1020. doi: 10.1111/nph.19446. Epub 2023 Dec 10. New Phytol. 2024. PMID: 38073141 Review.
-
Lessons in Effector and NLR Biology of Plant-Microbe Systems.Mol Plant Microbe Interact. 2018 Jan;31(1):34-45. doi: 10.1094/MPMI-08-17-0196-FI. Epub 2017 Nov 16. Mol Plant Microbe Interact. 2018. PMID: 29144205 Review.
Cited by
-
Status on Genetic Resistance to Rice Blast Disease in the Post-Genomic Era.Plants (Basel). 2025 Mar 5;14(5):807. doi: 10.3390/plants14050807. Plants (Basel). 2025. PMID: 40094775 Free PMC article. Review.
-
Zinc-finger (ZiF) fold secreted effectors form a functionally diverse family across lineages of the blast fungus Magnaporthe oryzae.PLoS Pathog. 2024 Jun 17;20(6):e1012277. doi: 10.1371/journal.ppat.1012277. eCollection 2024 Jun. PLoS Pathog. 2024. PMID: 38885263 Free PMC article.
-
The role of Exo70s in plant defense against pathogens and insect pests and their application for crop breeding.Mol Breed. 2025 Jan 21;45(2):17. doi: 10.1007/s11032-025-01539-3. eCollection 2025 Feb. Mol Breed. 2025. PMID: 39850652 Review.
-
The exocyst in context.Biochem Soc Trans. 2024 Oct 30;52(5):2113-2122. doi: 10.1042/BST20231401. Biochem Soc Trans. 2024. PMID: 39377315 Free PMC article. Review.
-
A glance at structural biology in advancing rice blast fungus research.Virulence. 2024 Dec;15(1):2403566. doi: 10.1080/21505594.2024.2403566. Epub 2024 Sep 16. Virulence. 2024. PMID: 39285518 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
Grants and funding
- BBS/E/J/000CA523/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- BB/V015508/1/UKRI | Biotechnology and Biological Sciences Research Council (BBSRC)
- BB/P012574/UKRI | Biotechnology and Biological Sciences Research Council (BBSRC)
- BBS/E/J/000PR9777/UKRI | Biotechnology and Biological Sciences Research Council (BBSRC)
- BBS/E/J/000PR9795/UKRI | Biotechnology and Biological Sciences Research Council (BBSRC)
LinkOut - more resources
Full Text Sources
Research Materials
