

SYK coordinates neuroprotective microglial responses in neurodegenerative disease
- PMID: 36257314
- PMCID: PMC9617784
- DOI: 10.1016/j.cell.2022.09.030
SYK coordinates neuroprotective microglial responses in neurodegenerative disease
Abstract
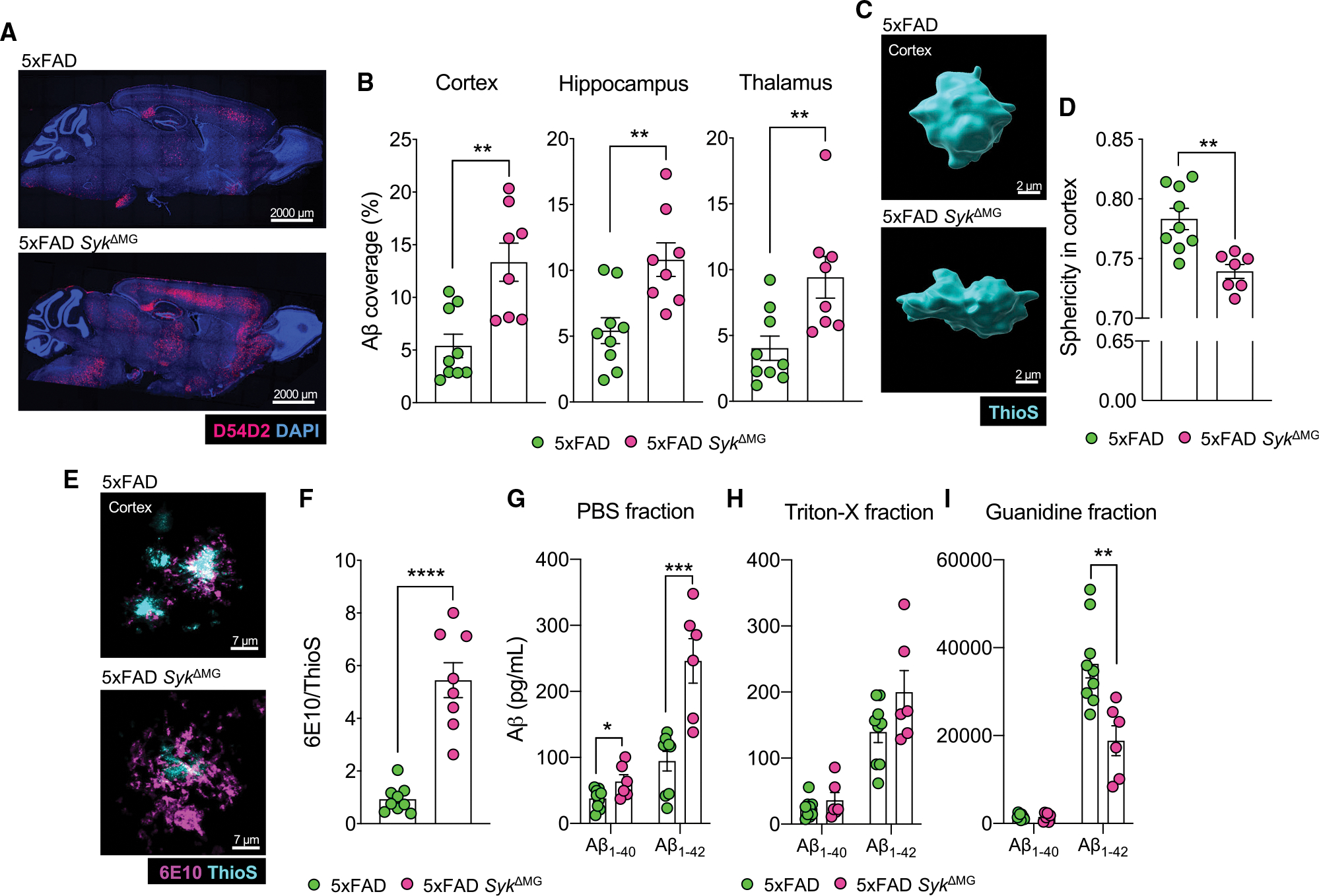
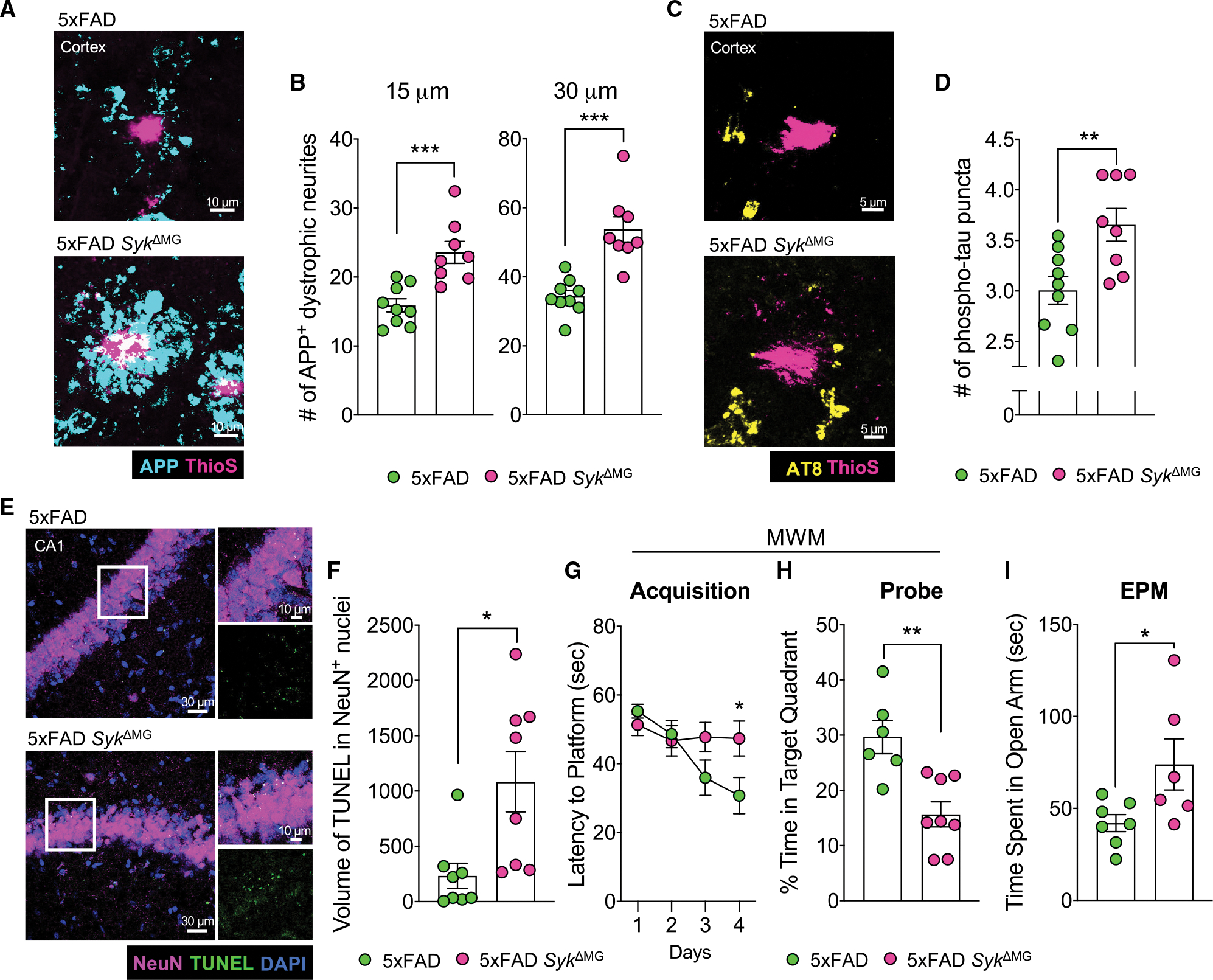
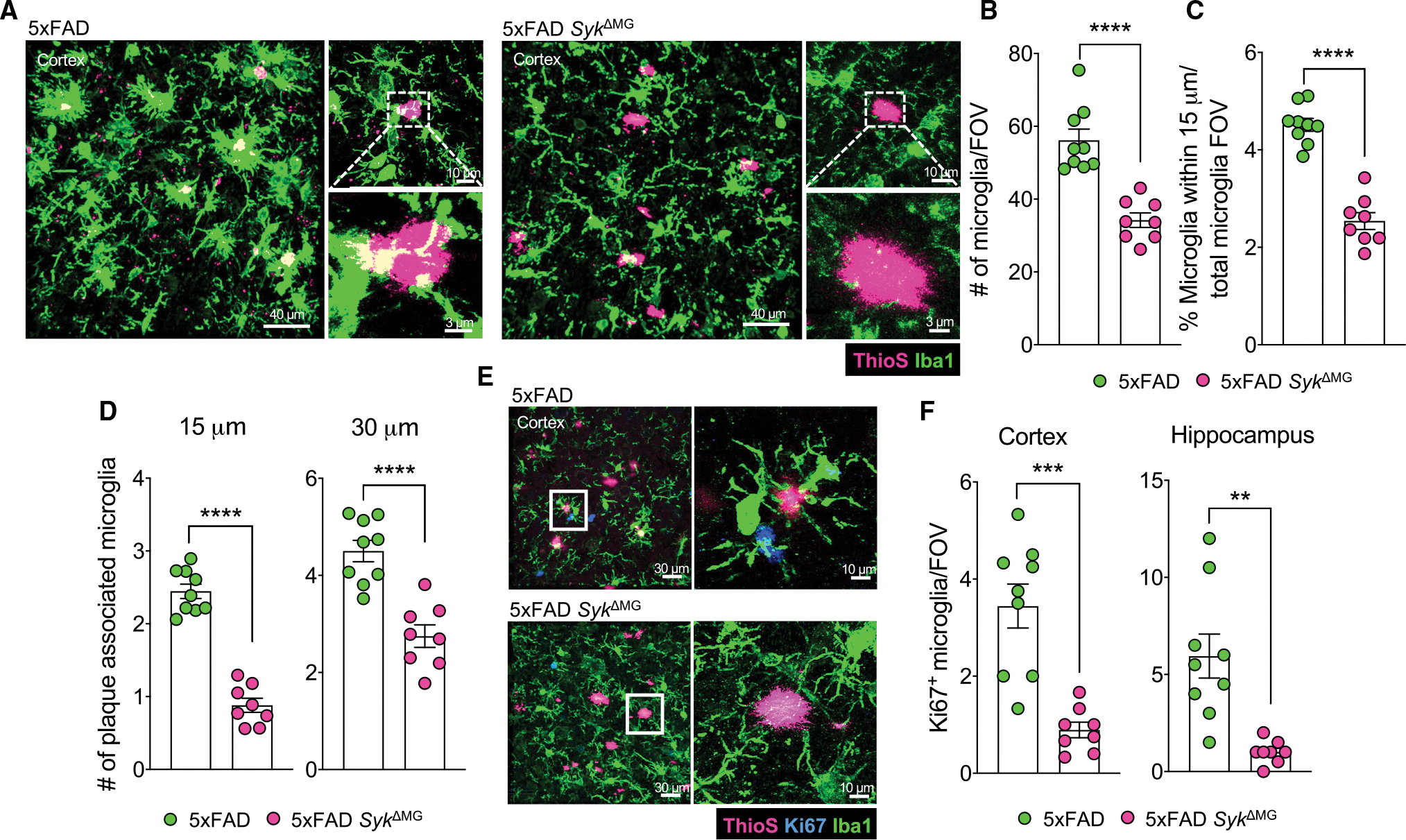
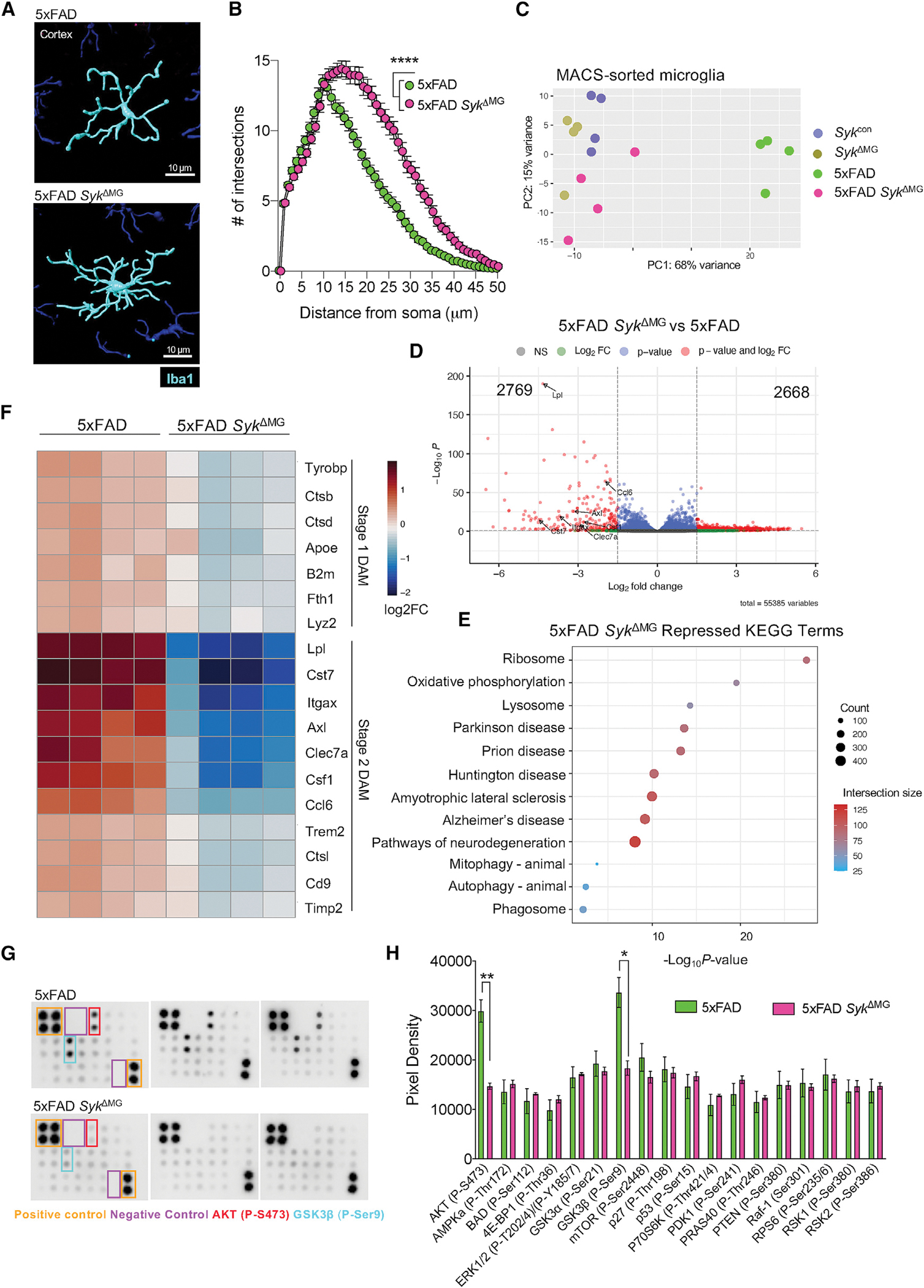
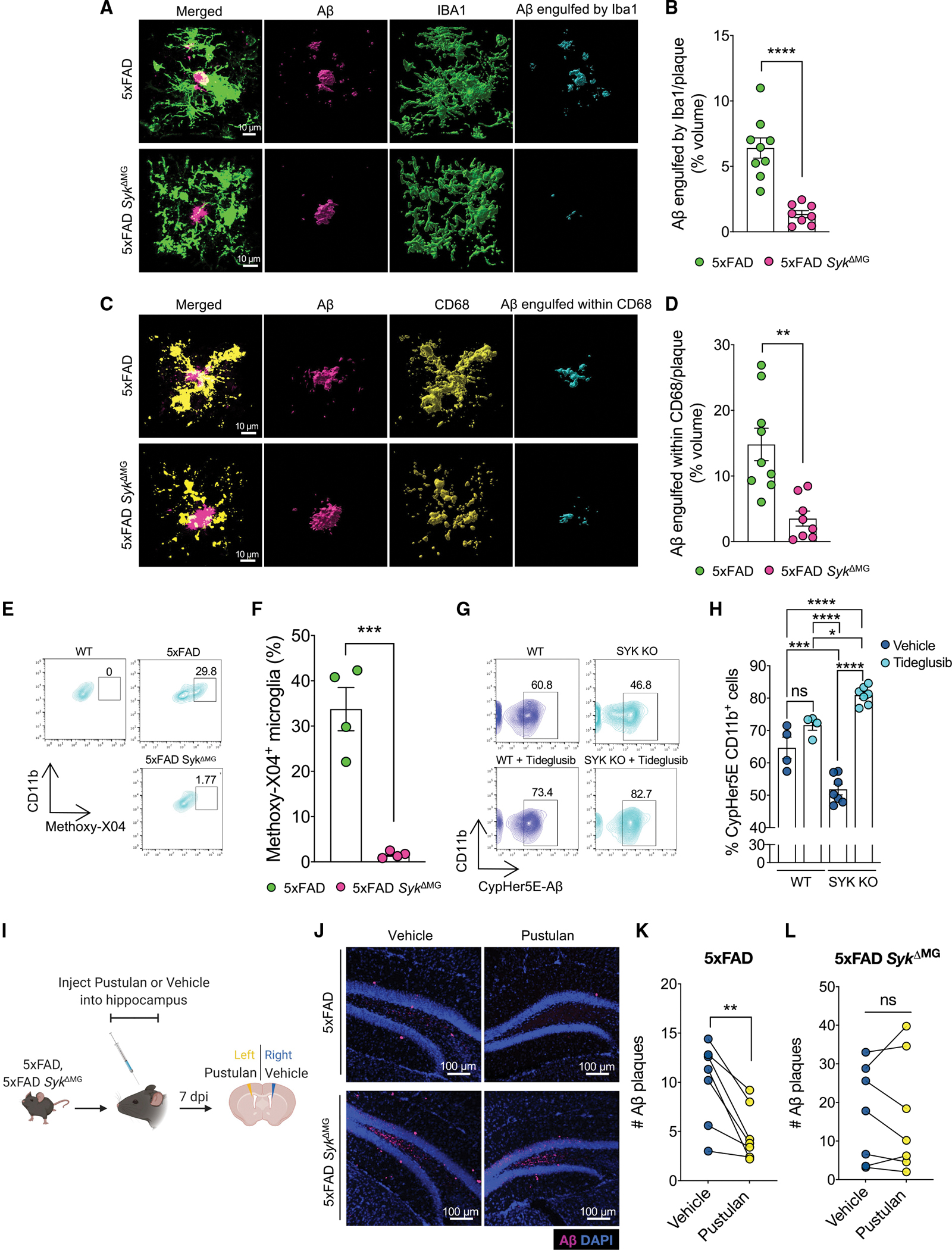
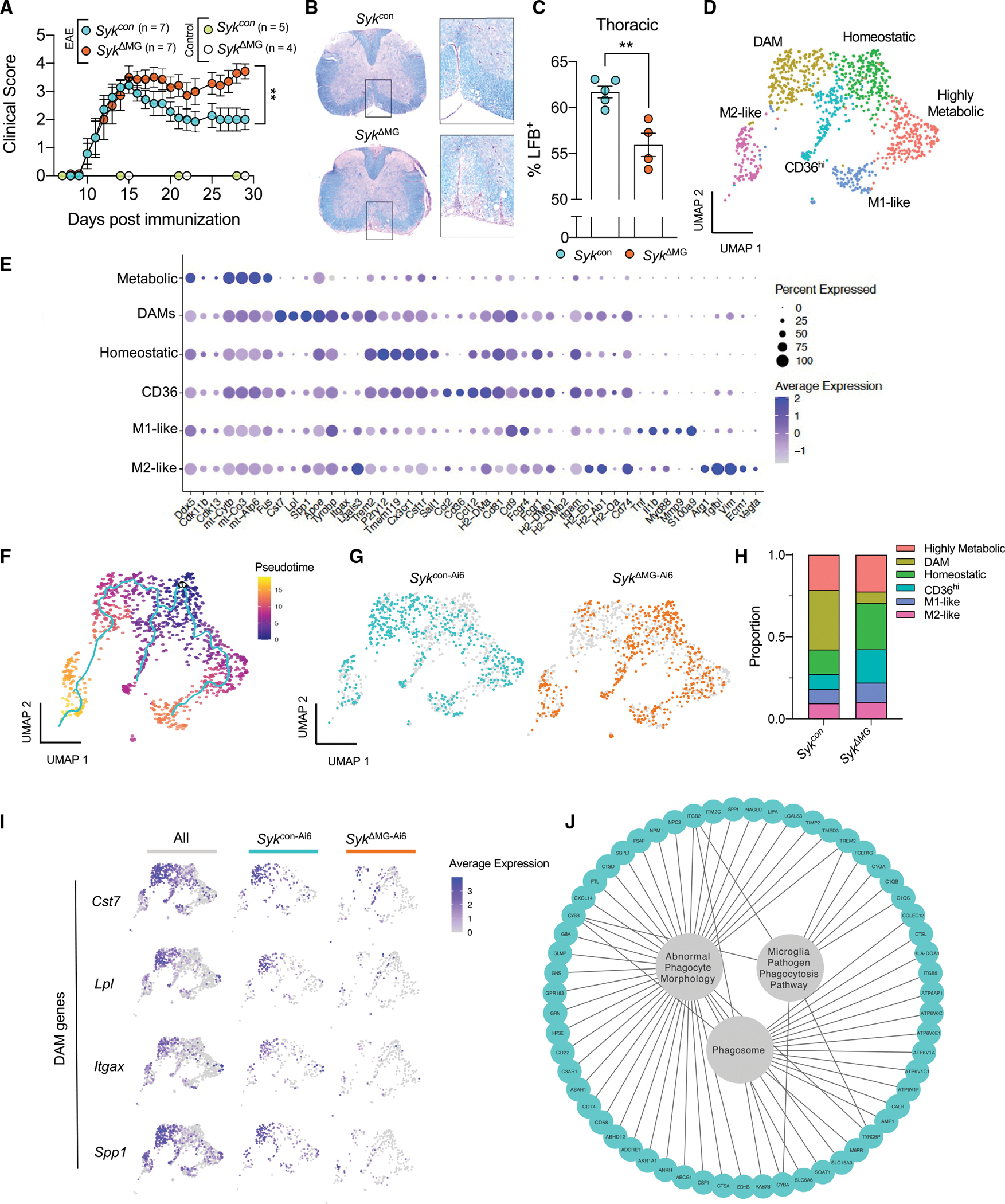
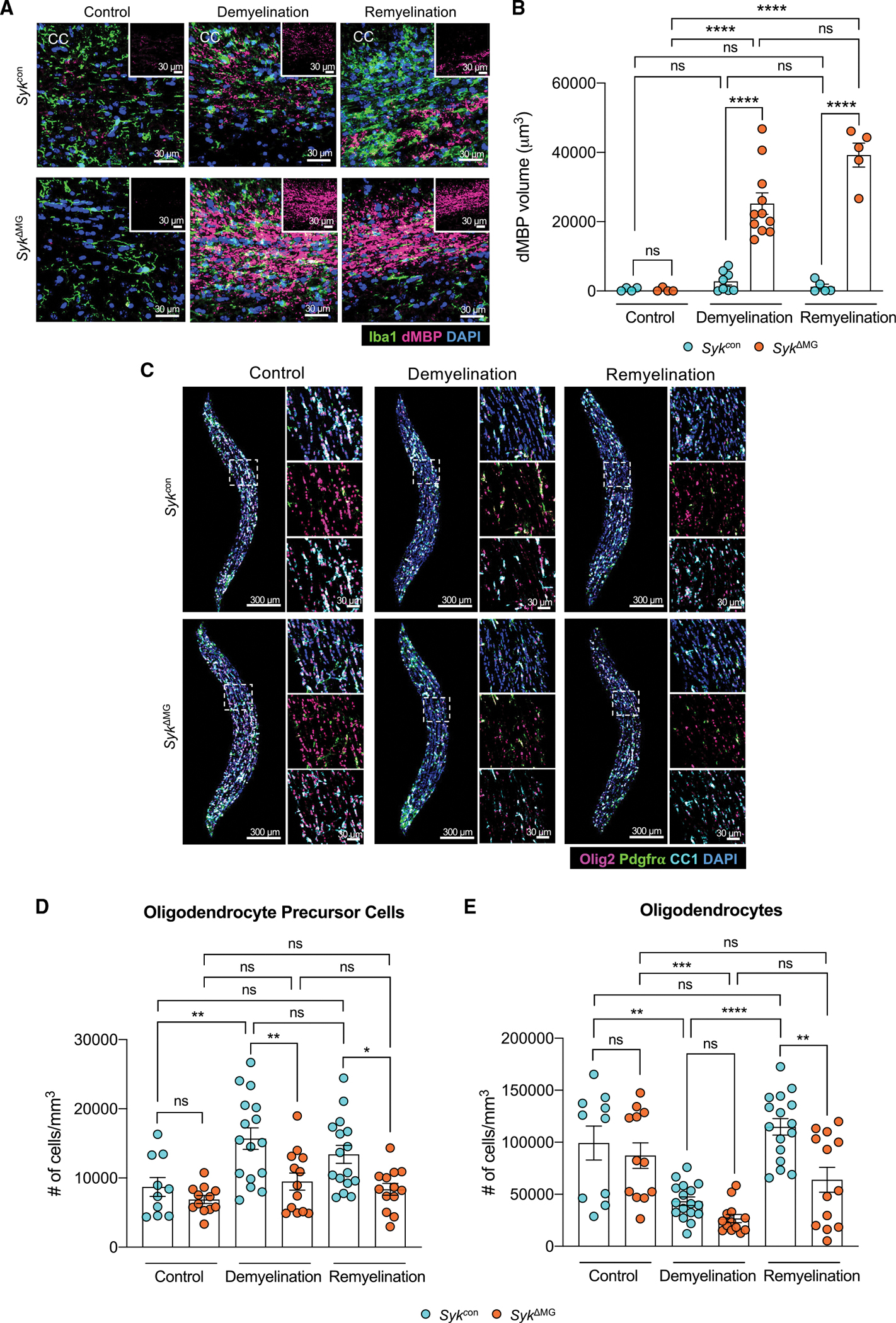
Recent studies have begun to reveal critical roles for the brain's professional phagocytes, microglia, and their receptors in the control of neurotoxic amyloid beta (Aβ) and myelin debris accumulation in neurodegenerative disease. However, the critical intracellular molecules that orchestrate neuroprotective functions of microglia remain poorly understood. In our studies, we find that targeted deletion of SYK in microglia leads to exacerbated Aβ deposition, aggravated neuropathology, and cognitive defects in the 5xFAD mouse model of Alzheimer's disease (AD). Disruption of SYK signaling in this AD model was further shown to impede the development of disease-associated microglia (DAM), alter AKT/GSK3β-signaling, and restrict Aβ phagocytosis by microglia. Conversely, receptor-mediated activation of SYK limits Aβ load. We also found that SYK critically regulates microglial phagocytosis and DAM acquisition in demyelinating disease. Collectively, these results broaden our understanding of the key innate immune signaling molecules that instruct beneficial microglial functions in response to neurotoxic material.
Keywords: Alzheimer’s disease; SYK; amyloid beta; disease-associated microglia; experimental autoimmune encephalomyelitis; microglia; multiple sclerosis; neurodegenerative disease; neuroimmunology; phagocytosis.
Copyright © 2022 The Author(s). Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
Microglia are SYK of Aβ and cell debris.Cell. 2022 Oct 27;185(22):4043-4045. doi: 10.1016/j.cell.2022.09.043. Cell. 2022. PMID: 36306731
References
-
- Bemiller SM, McCray TJ, Allan K, Formica SV, Xu G, Wilson G, Kokiko-Cochran ON, Crish SD, Lasagna-Reeves CA, Ransohoff RM, et al. (2017). TREM2 deficiency exacerbates tau pathology through dysregulated kinase signaling in a mouse model of tauopathy. Mol. Neurodegener. 12, 74. 10.1186/s13024-017-0216-6. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
