

The Essential Role of Water Molecules in the Reaction Mechanism of Protein O-Fucosyltransferase 2
- PMID: 36260536
- PMCID: PMC9828666
- DOI: 10.1002/anie.202213610
The Essential Role of Water Molecules in the Reaction Mechanism of Protein O-Fucosyltransferase 2
Abstract
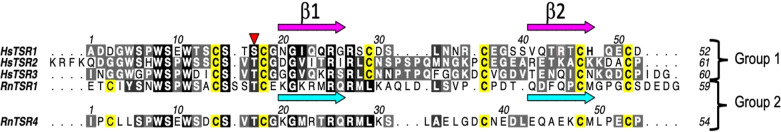
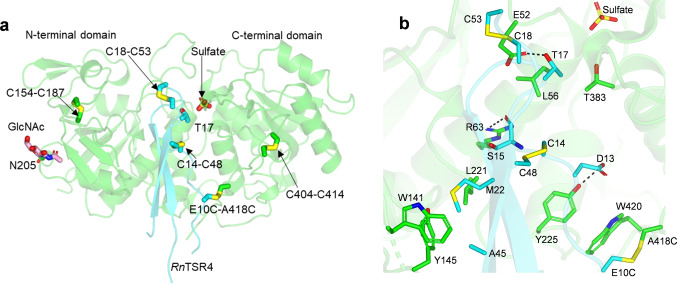
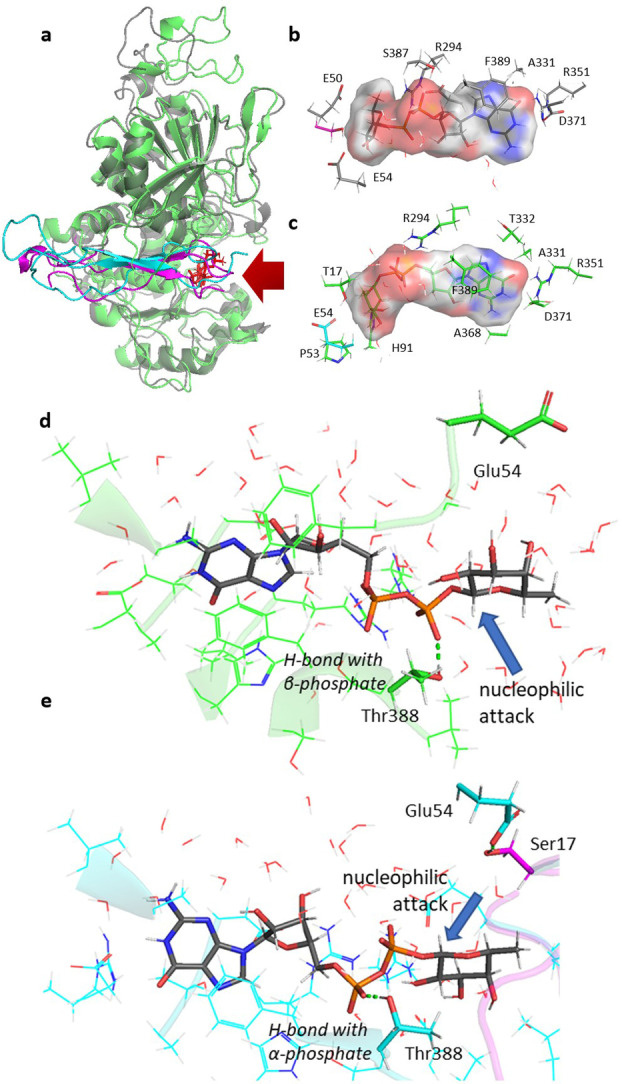
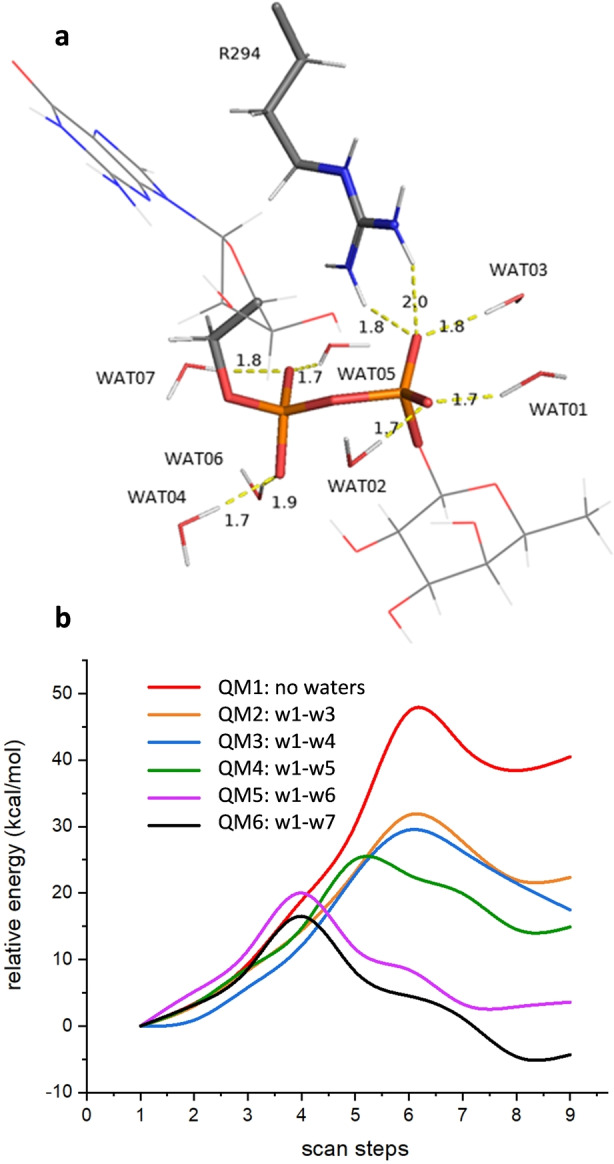
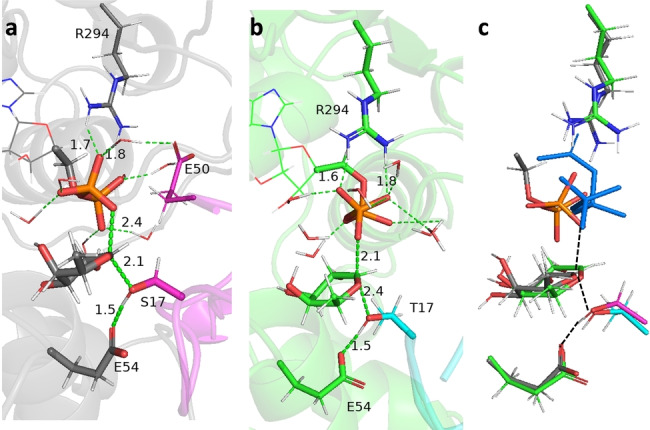
Protein O-fucosyltransferase 2 (PoFUT2) is an inverting glycosyltransferase (GT) that fucosylates thrombospondin repeats (TSRs) from group 1 and 2. PoFUT2 recognizes a large and diverse number of TSRs through a dynamic network of water-mediated interactions. By X-ray structural studies of C. elegans PoFUT2 complexed to a TSR of group 2, we demonstrate that this GT recognizes similarly the 3D structure of TSRs from both groups 1 and 2. Its active site is highly exposed to the solvent, suggesting that water molecules might also play an essential role in the fucosylation mechanism. We applied QM/MM methods using human PoFUT2 as a model, and found that HsPoFUT2 follows a classical SN 2 reaction mechanism in which water molecules contribute to a great extent in facilitating the release of the leaving pyrophosphate unit, causing the H transfer from the acceptor nucleophile (Thr/Ser) to the catalytic base, which is the last event in the reaction. This demonstrates the importance of water molecules not only in recognition of the ligands but also in catalysis.
Keywords: Enzyme Catalysis; Fucosylation; Glycosyltransferases; Protein Glycosylation; QM/MM.
© 2022 The Authors. Angewandte Chemie International Edition published by Wiley-VCH GmbH.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
A proactive role of water molecules in acceptor recognition by protein O-fucosyltransferase 2.Nat Chem Biol. 2016 Apr;12(4):240-6. doi: 10.1038/nchembio.2019. Epub 2016 Feb 8. Nat Chem Biol. 2016. PMID: 26854667 Free PMC article.
-
Structure of human POFUT2: insights into thrombospondin type 1 repeat fold and O-fucosylation.EMBO J. 2012 Jul 18;31(14):3183-97. doi: 10.1038/emboj.2012.143. EMBO J. 2012. PMID: 22588082 Free PMC article.
-
6-alkynyl fucose is a bioorthogonal analog for O-fucosylation of epidermal growth factor-like repeats and thrombospondin type-1 repeats by protein O-fucosyltransferases 1 and 2.Glycobiology. 2013 Feb;23(2):188-98. doi: 10.1093/glycob/cws140. Epub 2012 Oct 8. Glycobiology. 2013. PMID: 23045360 Free PMC article.
-
A perspective on structural and mechanistic aspects of protein O-fucosylation.Acta Crystallogr F Struct Biol Commun. 2018 Aug 1;74(Pt 8):443-450. doi: 10.1107/S2053230X18004788. Epub 2018 Jul 26. Acta Crystallogr F Struct Biol Commun. 2018. PMID: 30084393 Free PMC article. Review.
-
Protein O-fucosylation: structure and function.Curr Opin Struct Biol. 2019 Jun;56:78-86. doi: 10.1016/j.sbi.2018.12.005. Epub 2019 Jan 26. Curr Opin Struct Biol. 2019. PMID: 30690220 Free PMC article. Review.
Cited by
-
Reprogramming the Transition States to Enhance C-N Cleavage Efficiency of Rhodococcus opacusl-Amino Acid Oxidase.JACS Au. 2024 Jan 16;4(2):557-569. doi: 10.1021/jacsau.3c00672. eCollection 2024 Feb 26. JACS Au. 2024. PMID: 38425913 Free PMC article.
-
Aberrant glycosylation in schizophrenia: insights into pathophysiological mechanisms and therapeutic potentials.Front Pharmacol. 2024 Sep 2;15:1457811. doi: 10.3389/fphar.2024.1457811. eCollection 2024. Front Pharmacol. 2024. PMID: 39286629 Free PMC article. Review.
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
