

The E3 ligase adapter cereblon targets the C-terminal cyclic imide degron
- PMID: 36261529
- PMCID: PMC10316063
- DOI: 10.1038/s41586-022-05333-5
The E3 ligase adapter cereblon targets the C-terminal cyclic imide degron
Abstract
The ubiquitin E3 ligase substrate adapter cereblon (CRBN) is a target of thalidomide and lenalidomide1, therapeutic agents used in the treatment of haematopoietic malignancies2-4 and as ligands for targeted protein degradation5-7. These agents are proposed to mimic a naturally occurring degron; however, the structural motif recognized by the thalidomide-binding domain of CRBN remains unknown. Here we report that C-terminal cyclic imides, post-translational modifications that arise from intramolecular cyclization of glutamine or asparagine residues, are physiological degrons on substrates for CRBN. Dipeptides bearing the C-terminal cyclic imide degron substitute for thalidomide when embedded within bifunctional chemical degraders. Addition of the degron to the C terminus of proteins induces CRBN-dependent ubiquitination and degradation in vitro and in cells. C-terminal cyclic imides form adventitiously on physiologically relevant timescales throughout the human proteome to afford a degron that is endogenously recognized and removed by CRBN. The discovery of the C-terminal cyclic imide degron defines a regulatory process that may affect the physiological function and therapeutic engagement of CRBN.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Figures





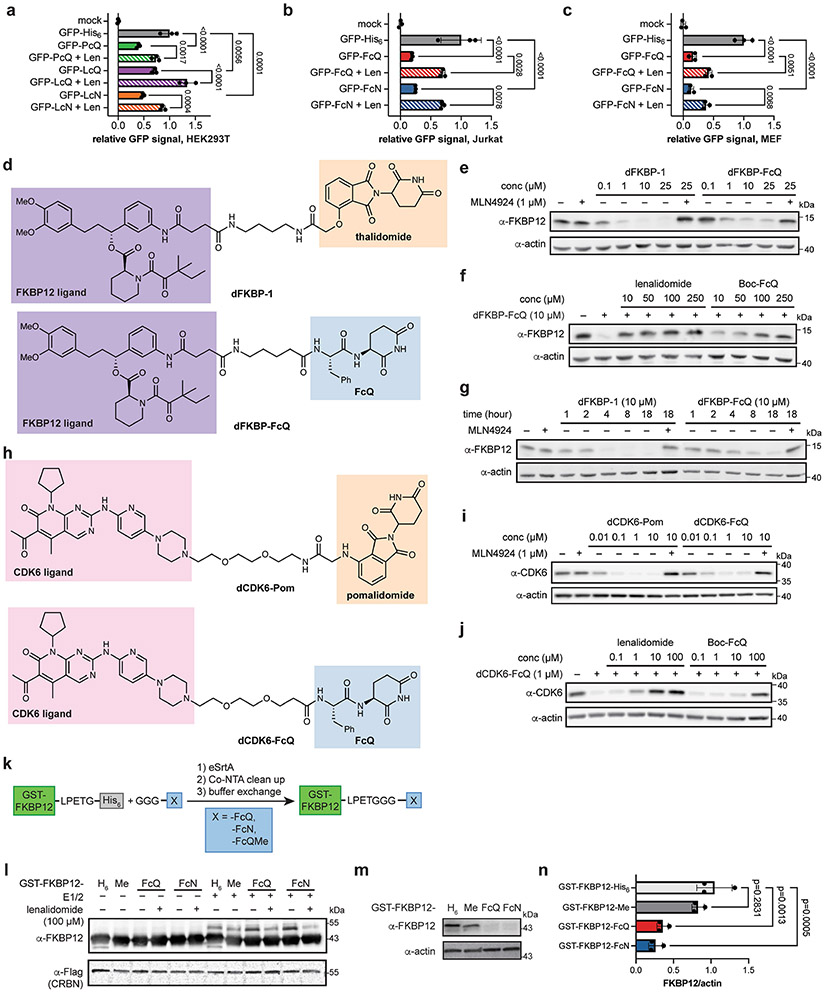
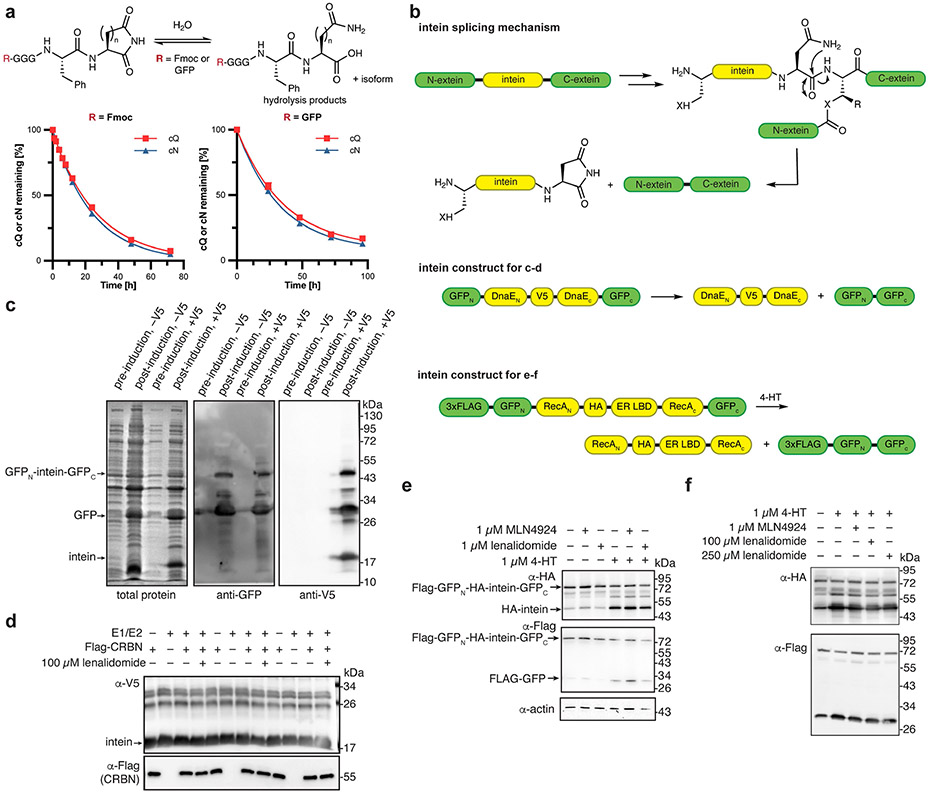

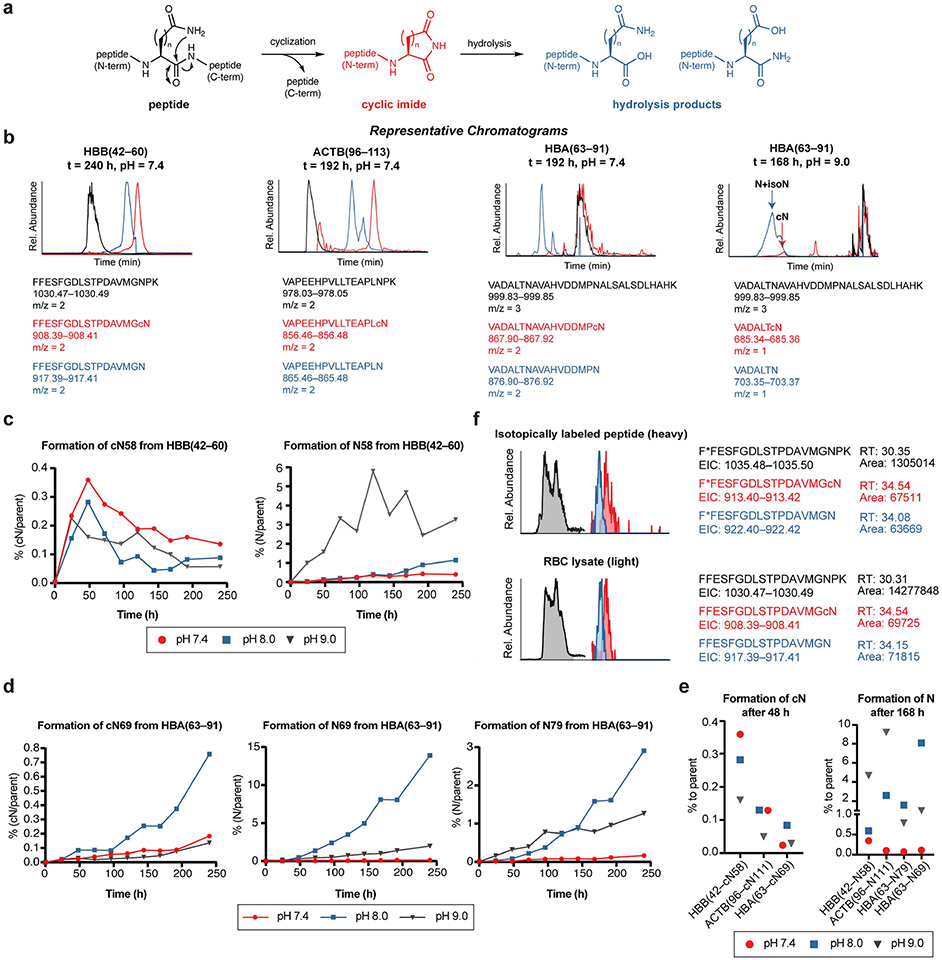


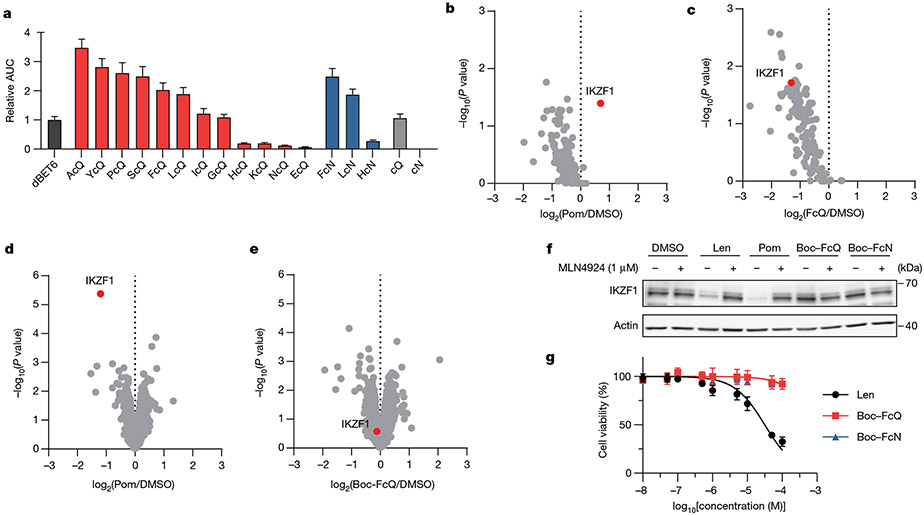



Comment in
-
A closer look at molecular glues.Nat Rev Drug Discov. 2022 Dec;21(12):880. doi: 10.1038/d41573-022-00184-1. Nat Rev Drug Discov. 2022. PMID: 36323853 No abstract available.
References
-
- Ito T et al. Identification of a primary target of thalidomide teratogenicity. Science 327, 1345–1350 (2010). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
