

Time-restricted feeding mitigates obesity through adipocyte thermogenesis
- PMID: 36264811
- PMCID: PMC10150371
- DOI: 10.1126/science.abl8007
Time-restricted feeding mitigates obesity through adipocyte thermogenesis
Abstract
Misalignment of feeding rhythms with the light-dark cycle leads to disrupted peripheral circadian clocks and obesity. Conversely, restricting feeding to the active period mitigates metabolic syndrome through mechanisms that remain unknown. We found that genetic enhancement of adipocyte thermogenesis through ablation of the zinc finger protein 423 (ZFP423) attenuated obesity caused by consumption of a high-fat diet during the inactive (light) period by increasing futile creatine cycling in mice. Circadian control of adipocyte creatine metabolism underlies the timing of diet-induced thermogenesis, and enhancement of adipocyte circadian rhythms through overexpression of the clock activator brain and muscle Arnt-like protein-1 (BMAL1) ameliorated metabolic complications during diet-induced obesity. These findings uncover rhythmic creatine-mediated thermogenesis as an essential mechanism that drives metabolic benefits during time-restricted feeding.
Conflict of interest statement
Figures
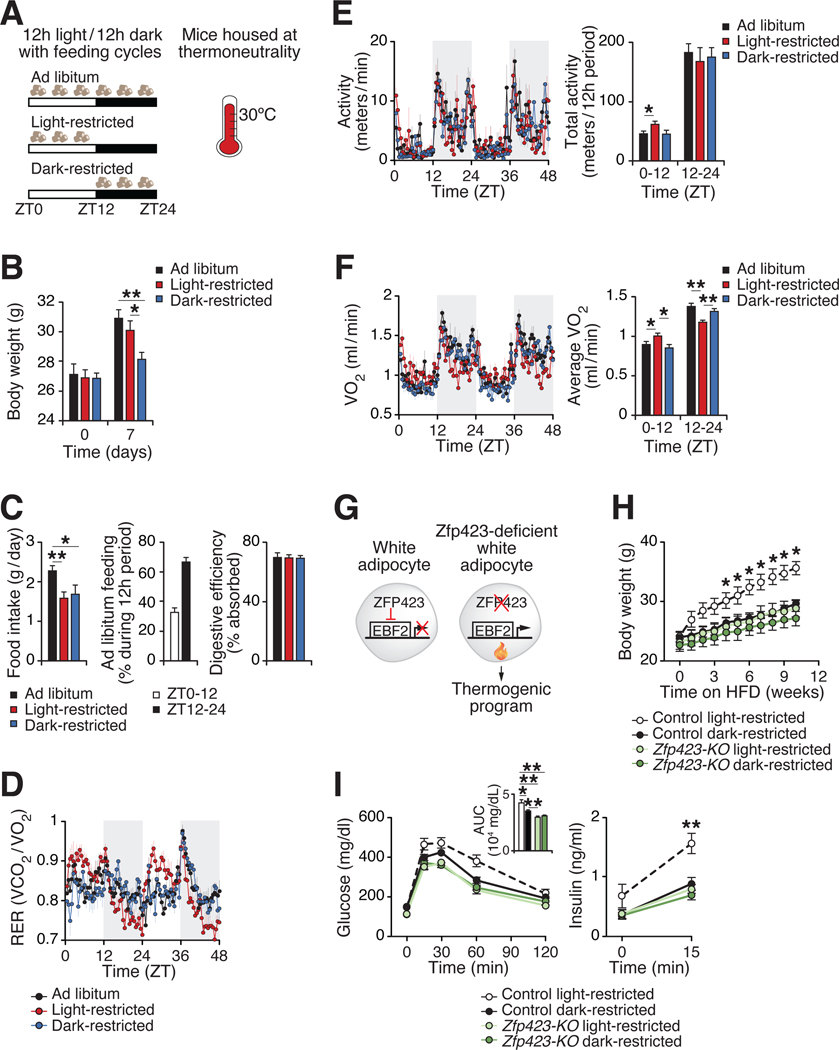


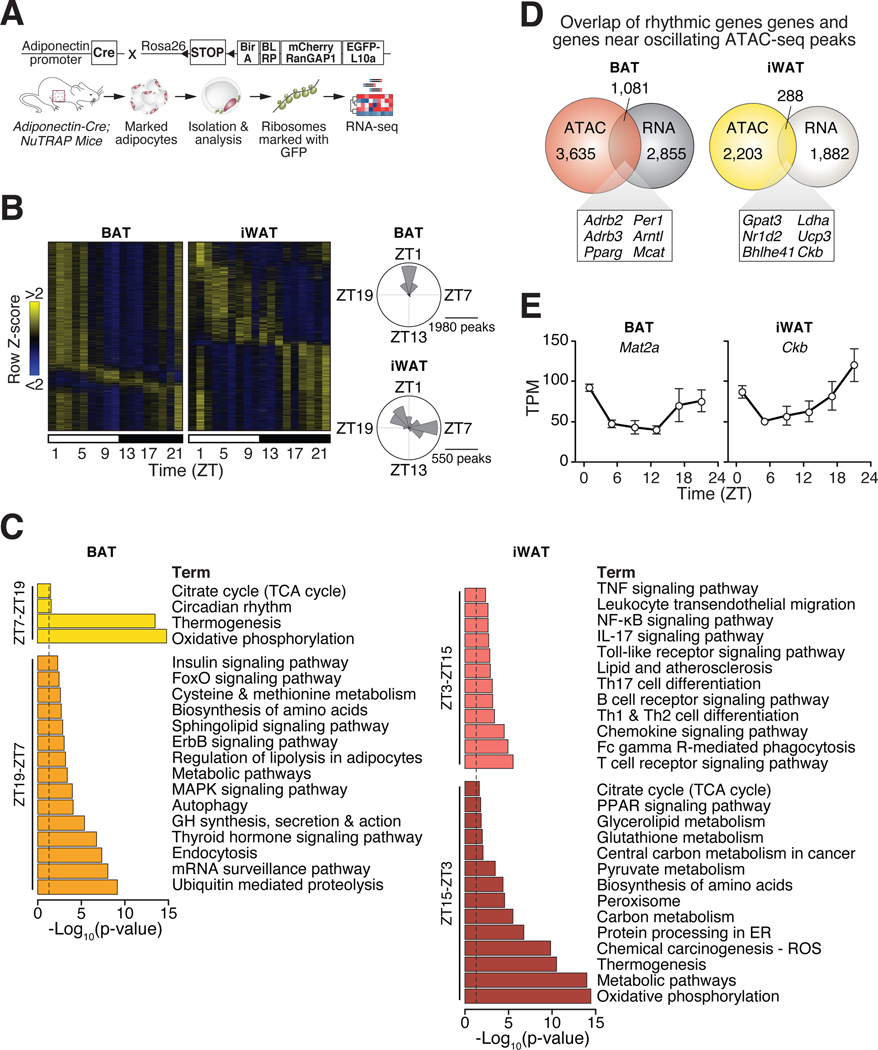

Comment in
-
The timing of eating controls energy use.Science. 2022 Oct 21;378(6617):251-252. doi: 10.1126/science.ade6720. Epub 2022 Oct 20. Science. 2022. PMID: 36264788
-
Timed feeding aligns the adipocyte clock to optimize thermogenesis when eating a high-fat diet.Cell Metab. 2023 Jan 3;35(1):7-9. doi: 10.1016/j.cmet.2022.12.008. Cell Metab. 2023. PMID: 36599303 Free PMC article.
-
Timing of food intake affects energy expenditure.Nat Rev Endocrinol. 2023 Oct;19(10):559. doi: 10.1038/s41574-023-00878-5. Nat Rev Endocrinol. 2023. PMID: 37488263 No abstract available.
References
-
- Kohsaka A. et al. , High-fat diet disrupts behavioral and molecular circadian rhythms in mice. Cell Metab 6, 414–421 (2007). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- F31 DK130589/DK/NIDDK NIH HHS/United States
- R01 DK119163/DK/NIDDK NIH HHS/United States
- R01 DK090625/DK/NIDDK NIH HHS/United States
- F30 DK116481/DK/NIDDK NIH HHS/United States
- R01 DK104789/DK/NIDDK NIH HHS/United States
- K99 DK124682/DK/NIDDK NIH HHS/United States
- R01 AG065988/AG/NIA NIH HHS/United States
- P01 AG011412/AG/NIA NIH HHS/United States
- R01 DK113011/DK/NIDDK NIH HHS/United States
- F32 DK122675/DK/NIDDK NIH HHS/United States
- R01 DK127800/DK/NIDDK NIH HHS/United States
- R01 DK050203/DK/NIDDK NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
