

Single-molecule dynamics of surface lipoproteins in bacteroides indicate similarities and cooperativity
- PMID: 36266970
- PMCID: PMC9748367
- DOI: 10.1016/j.bpj.2022.10.024
Single-molecule dynamics of surface lipoproteins in bacteroides indicate similarities and cooperativity
Abstract
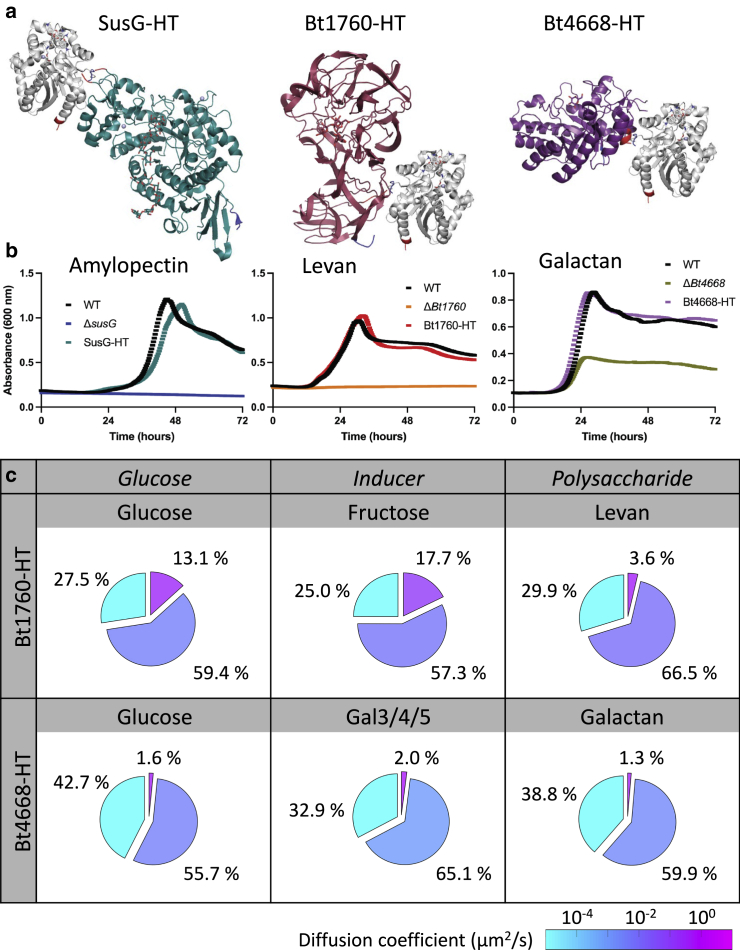
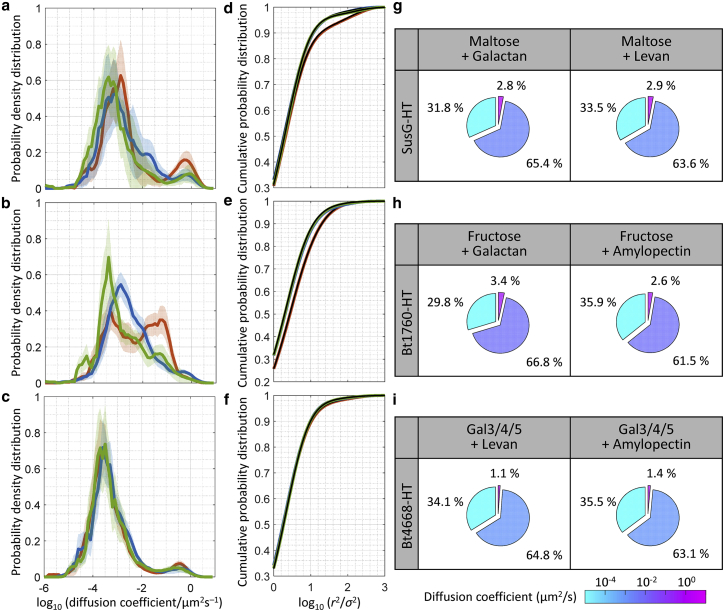
The gut microbiota comprises hundreds of species with a composition shaped by the available glycans. The well-studied starch utilization system (Sus) is a prototype for glycan uptake in the human gut bacterium Bacteroides thetaiotaomicron (Bt). Each Sus-like system includes outer-membrane proteins, which translocate glycan into the periplasm, and one or more cell-surface glycoside hydrolases, which break down a specific (cognate) polymer substrate. Although the molecular mechanisms of the Sus system are known, how the Sus and Sus-like proteins cooperate remains elusive. Previously, we used single-molecule and super-resolution fluorescence microscopy to show that SusG is mobile on the outer membrane and slows down in the presence of starch. Here, we compare the dynamics of three glycoside hydrolases: SusG, Bt4668, and Bt1760, which target starch, galactan, and levan, respectively. We characterized the diffusion of each surface hydrolase in the presence of its cognate glycan and found that all three enzymes are mostly immobile in the presence of the polysaccharide, consistent with carbohydrate binding. Moreover, experiments in glucose versus oligosaccharides suggest that the enzyme dynamics depend on their expression level. Furthermore, we characterized enzyme diffusion in a mixture of glycans and found that noncognate polysaccharides modify the dynamics of SusG and Bt1760 but not Bt4668. We investigated these systems with polysaccharide mixtures and genetic knockouts and found that noncognate polysaccharides modify hydrolase dynamics through some combination of nonspecific protein interactions and downregulation of the hydrolase. Overall, these experiments extend our understanding of how Sus-like lipoprotein dynamics can be modified by changing carbohydrate conditions and the expression level of the enzyme.
Copyright © 2022 Biophysical Society. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures




References
-
- Ndeh D., Gilbert H.J. Biochemistry of complex glycan depolymerisation by the human gut microbiota. FEMS Microbiol. Rev. 2018;42:146–164. - PubMed
-
- Makki K., Deehan E.C., et al. Bäckhed F. The impact of dietary fiber on gut microbiota in host health and disease. Cell Host Microbe. 2018;23:705–715. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
