

doi: 10.1093/plphys/kiac493.
Respiratory energy demands and scope for demand expansion and destruction
Affiliations
- PMID: 36271857
- PMCID: PMC10069906
- DOI: 10.1093/plphys/kiac493
Item in Clipboard
Respiratory energy demands and scope for demand expansion and destruction
Plant Physiol.
.
Abstract
Nonphotosynthetic plant metabolic processes are powered by respiratory energy, a limited resource that metabolic engineers—like plants themselves—must manage prudently.
Conflict of interest statement
Conflict of interest statement. None declared.
Figures

Plant respiratory metabolism and what it fuels. A, Overview of respiratory metabolism. B, The approximate fractions of total photosynthate that are respired to fuel growth or maintenance processes, or are allocated to harvested or nonharvested biomass. OPP pathway, oxidative pentose phosphate pathway; G6P, glucose 6-phosphate; Ox-Phos, oxidative phosphorylation; TCA cycle, tricarboxylic acid cycle.

Energy demands of growth processes and protein turnover. A and B, grams of hexose required to make 1 g of product at maximum theoretical efficiency of biosynthesis. A, Hexose costs, with ammonium or nitrate as N source, for three progressively more expensive amino acids. Biosynthetic pathway analysis from Amthor (2010), using 28 ATP/hexose, 11.5 NAD(P)H/hexose, and nitrate uptake and reduction costs as in Supplemental Appendix S1. B, Hexose costs of various crop storage organs: sugar beet roots, potato tubers, inflorescences with seeds (rice, wheat, and sunflower), maize cobs (70% seeds), pods with seeds (chickpea and soybean), and oil palm fruits. Carbohydrate, protein, lipid, lignin, organic acid, and mineral concentrations in organs are based on Penning de Vries et al. (1983). Substrate requirements for biosynthesis of each class of constituent are from Penning de Vries et al. (1983) and Amthor (2010) (for lignin and carbohydrates). Tool maintenance estimates are included, and 5% of the carbohydrate substrate is respired to provide energy to transport the other 95% from sources to storage organs. Minerals are supplied as needed, accounting for uptake cost in roots. The N source is nitrate and its assimilation cost is included. C, Hexose costs of breakdown plus resynthesis of eight Arabidopsis leaf proteins, recalculated from data of Li et al. (2017). DW, dry weight.
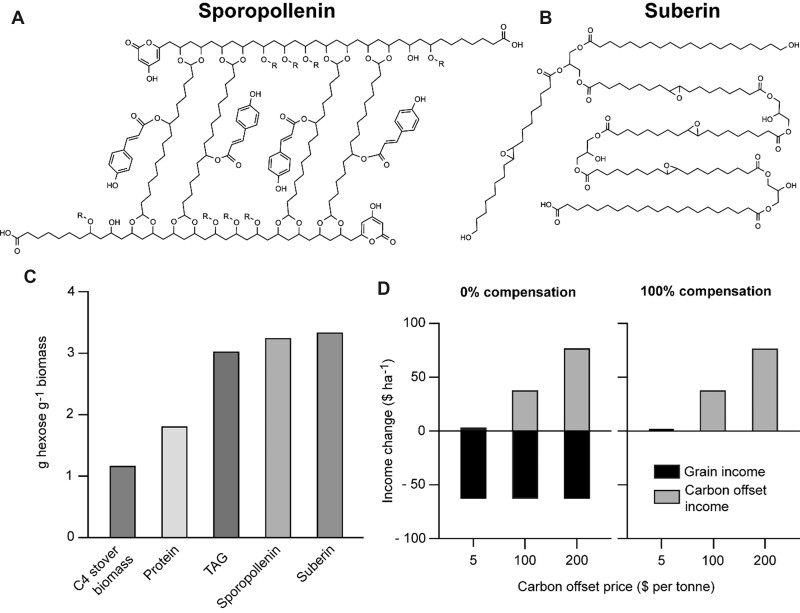
Potential root-synthesized, recalcitrant CCS polymers and their economics. A, The molecular structure of sporopollenin, based on pine sporopollenin analysis (Li et al., 2019). R is glycerol. B, The molecular structure of suberin (Pereira, 2015). C, Calculated hexose costs for synthesis of typical C4 stover biomass, protein, triacylglycerol (TAG, specifically triolein), sporopollenin, and suberin, based on Amthor’s values (2010) and Supplemental Table S2. D, Estimated impacts on farm income per hectare of engineering maize roots to make extra suberin or sporopollenin equal to 10% of root dry weight, in two scenarios (0% or 100% compensation of the added carbon and energy demand by increased photosynthesis or decreased respiration). Grain price is $166 per tonne; carbon offset price is $5, $100, or $200 per tonne of CO2.

Obtaining N from in planta BNF or diazotroph BNF instead of fertilizer nitrate. A, Potential sources of N for maize: 1, Soil nitrate (or ammonium) from fertilizer, taken up by roots; 2, Nitrogenase system engineered in leaves; 3, Soil diazotrophs that transfer fixed N to the plant via the roots. B, The hexose cost of N (assimilated into glutamate) when the source is soil nitrate, soil ammonium, or atmospheric N2. N2 costs are calculated for an engineered leaf nitrogenase (in planta nitrogenase) and for BNF by root-associated diazotrophs (diazotroph), and for low or high ATP requirements (16 or 40 ATP per N2, respectively). C, Estimated maize grain yields for crops in which BNF replaces all the N (120 kg ha−1) that a benchmark crop obtains from nitrate, and the amounts of photosynthate (as hexose) exuded from roots that would be needed to support diazotroph BNF. Calculations assume diazotrophs use either 100% or 25% of the exuded hexose for N fixation.

Predicted impact of reduced protein turnover cost on crop yield. Percentage gain in whole crop biomass carbon accumulation and grain carbon storage for a crop with 6.5% reduction in maintenance respiration relative to the unmodified crop. Grain carbon accumulation was initiated after anthesis during Day 200 of the year.
References
-
- Ainsworth EA, Long SP (2005) What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol 165: 351–371 - PubMed
-
- Amthor JS (1989) Respiration and Crop Productivity. Springer-Verlag, New York, NY
-
- Amthor JS (1994) Respiration and carbon assimilate use. InBoote KJ, Bennett JM, Sinclair TR, Paulsen GM, eds, Physiology and Determination of Crop Yield. American Society of Agronomy, Madison, WI, pp 221–250
-
- Amthor JS (2000) The McCree–de Wit–Penning de Vries–Thornley respiration paradigms: 30 years later. Ann Bot 86: 1–20
-
- Amthor JS (2010) From sunlight to phytomass: on the potential efficiency of converting solar radiation to phyto-energy. New Phytol 188: 939–959 - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
