

A bovine antibody possessing an ultralong complementarity-determining region CDRH3 targets a highly conserved epitope in sarbecovirus spike proteins
- PMID: 36272646
- PMCID: PMC9678781
- DOI: 10.1016/j.jbc.2022.102624
A bovine antibody possessing an ultralong complementarity-determining region CDRH3 targets a highly conserved epitope in sarbecovirus spike proteins
Abstract
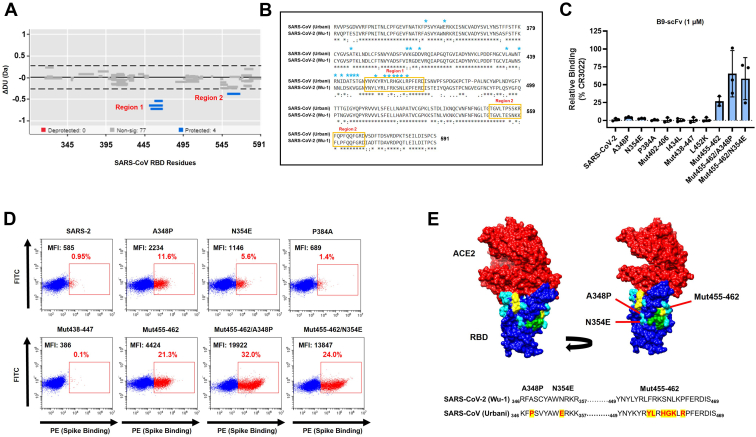
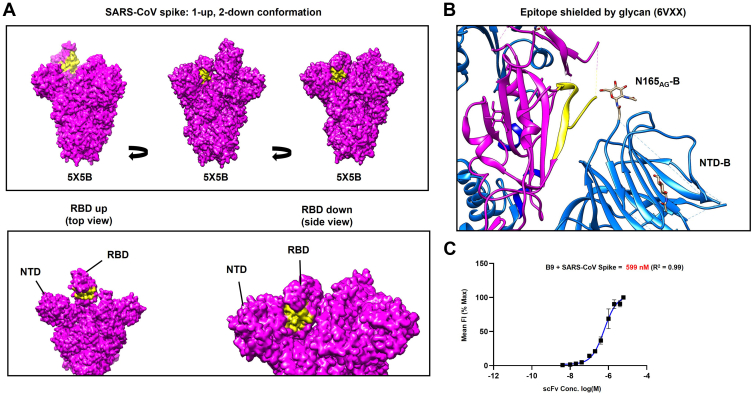
Broadly neutralizing antibodies have huge potential as novel antiviral therapeutics due to their ability to recognize highly conserved epitopes that are seldom mutated in viral variants. A subset of bovine antibodies possess an ultralong complementarity-determining region (CDR)H3 that is highly adept at recognizing such conserved epitopes, but their reactivity against Sarbecovirus Spike proteins has not been explored previously. Here, we use a SARS-naïve library to isolate a broadly reactive bovine CDRH3 that binds the receptor-binding domain of SARS-CoV, SARS-CoV-2, and all SARS-CoV-2 variants. We show further that it neutralizes viruses pseudo-typed with SARS-CoV Spike, but this is not by competition with angiotensin-converting enzyme 2 (ACE2) binding. Instead, using differential hydrogen-deuterium exchange mass spectrometry, we demonstrate that it recognizes the major site of vulnerability of Sarbecoviruses. This glycan-shielded cryptic epitope becomes available only transiently via interdomain movements of the Spike protein such that antibody binding triggers destruction of the prefusion complex. This proof of principle study demonstrates the power of in vitro expressed bovine antibodies with ultralong CDRH3s for the isolation of novel, broadly reactive tools to combat emerging pathogens and to identify key epitopes for vaccine development.
Keywords: antibody; antibody engineering; antiviral agent; epitope mapping; hydrogen-deuterium exchange.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflicts of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures
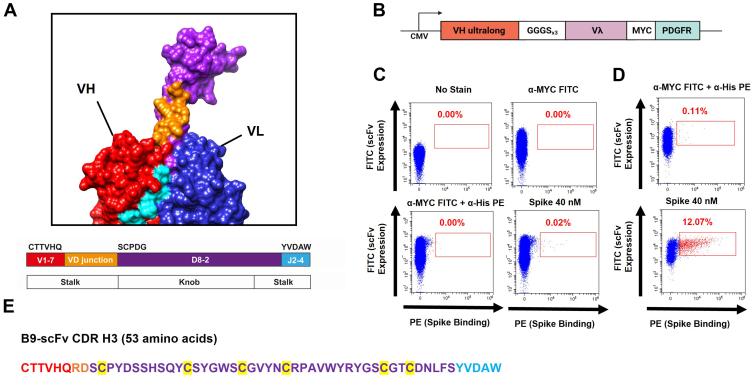
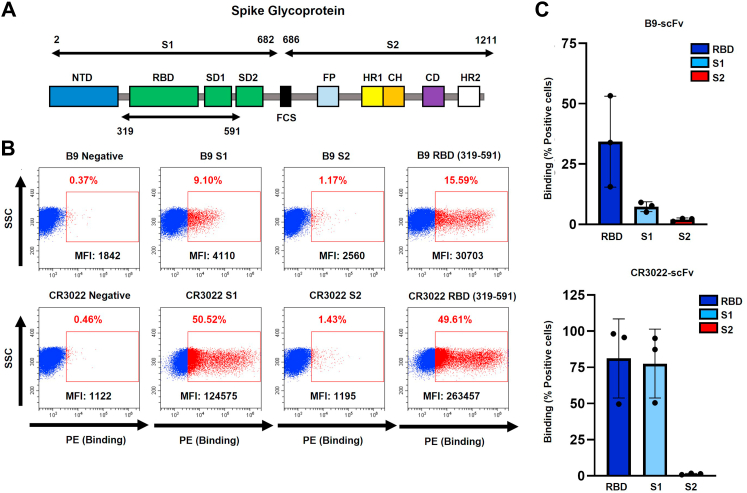
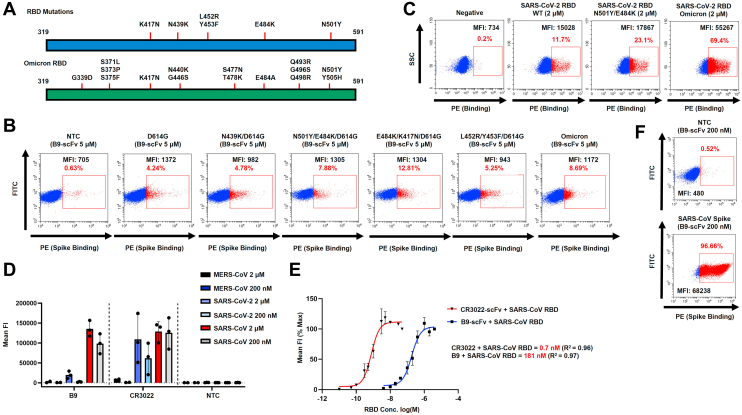
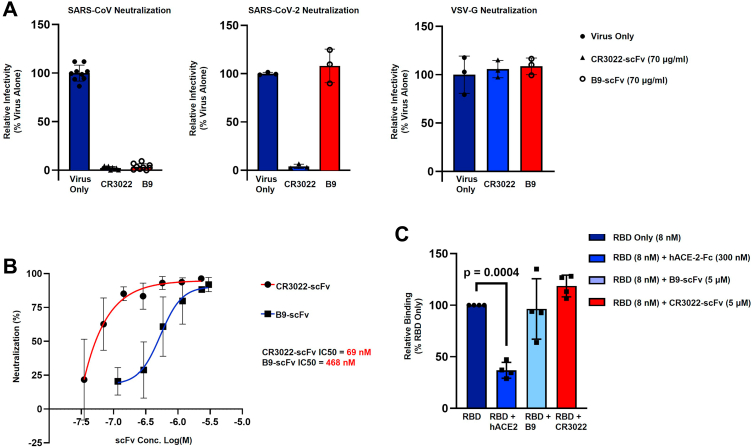
References
-
- Voysey M., Clemens S.A.C., Madhi S.A., Weckx L.Y., Folegatti P.M., Aley P.K., et al. Safety and efficacy of the ChAdOx1 nCoV-19 vaccine (AZD1222) against SARS-CoV-2: an interim analysis of four randomised controlled trials in Brazil, South Africa, and the UK. The Lancet. 2021;397:99–111. - PMC - PubMed
Publication types
MeSH terms
Substances
Supplementary concepts
Grants and funding
- BB/M012573/1/BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- 220628/Z/20/Z/WT_/Wellcome Trust/United Kingdom
- 220628/WT_/Wellcome Trust/United Kingdom
- BB/V01384X/1 /BB_/Biotechnology and Biological Sciences Research Council/United Kingdom
- MR/N013840/1/MRC_/Medical Research Council/United Kingdom
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
