

Genome-scale analysis of Arabidopsis splicing-related protein kinase families reveals roles in abiotic stress adaptation
- PMID: 36273172
- PMCID: PMC9587599
- DOI: 10.1186/s12870-022-03870-9
Genome-scale analysis of Arabidopsis splicing-related protein kinase families reveals roles in abiotic stress adaptation
Erratum in
-
Correction: Genome-scale analysis of Arabidopsis splicing-related protein kinase families reveals roles in abiotic stress adaptation.BMC Plant Biol. 2023 May 23;23(1):272. doi: 10.1186/s12870-023-04289-6. BMC Plant Biol. 2023. PMID: 37217886 Free PMC article. No abstract available.
Abstract
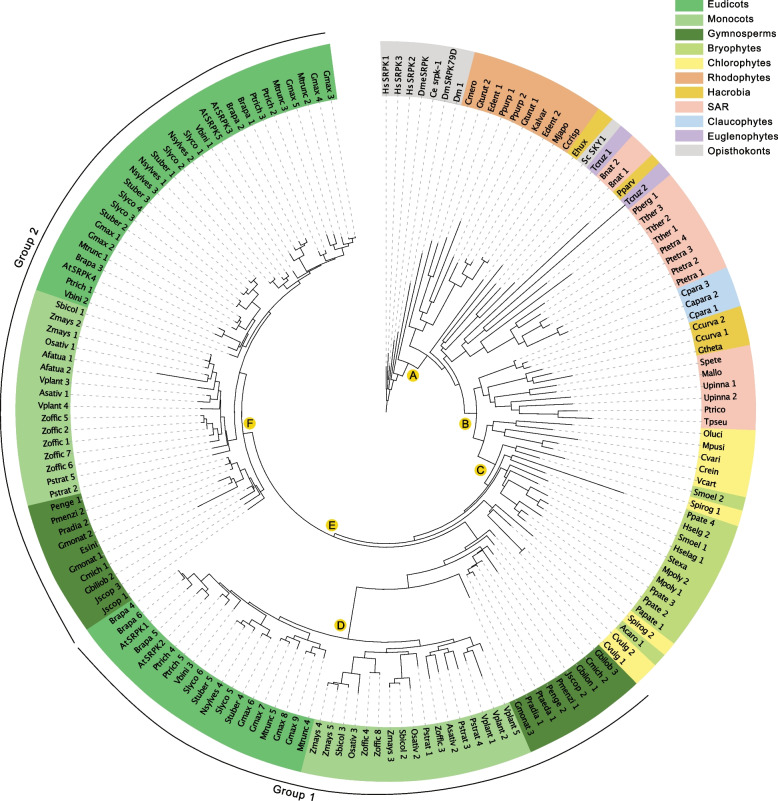
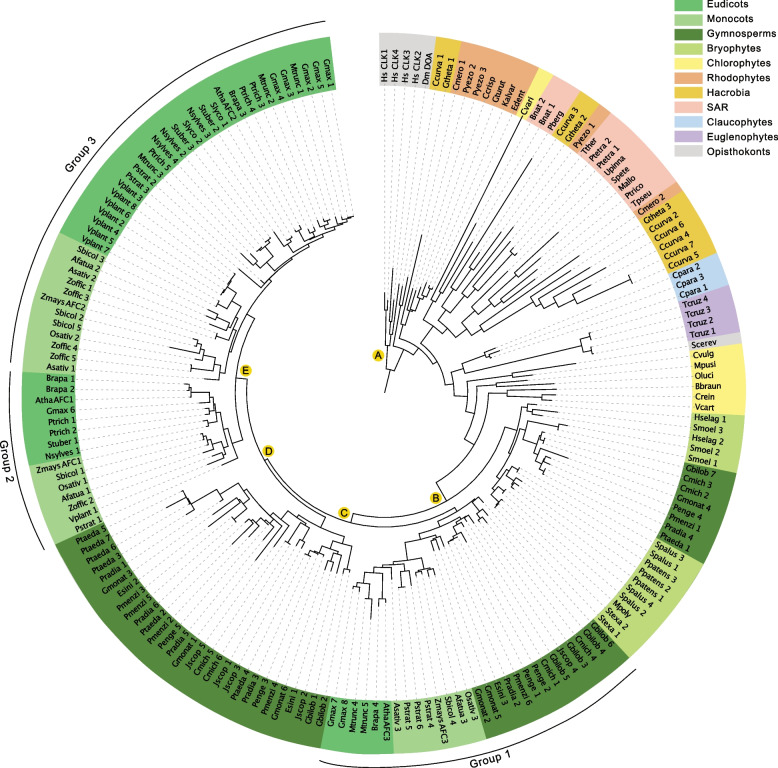
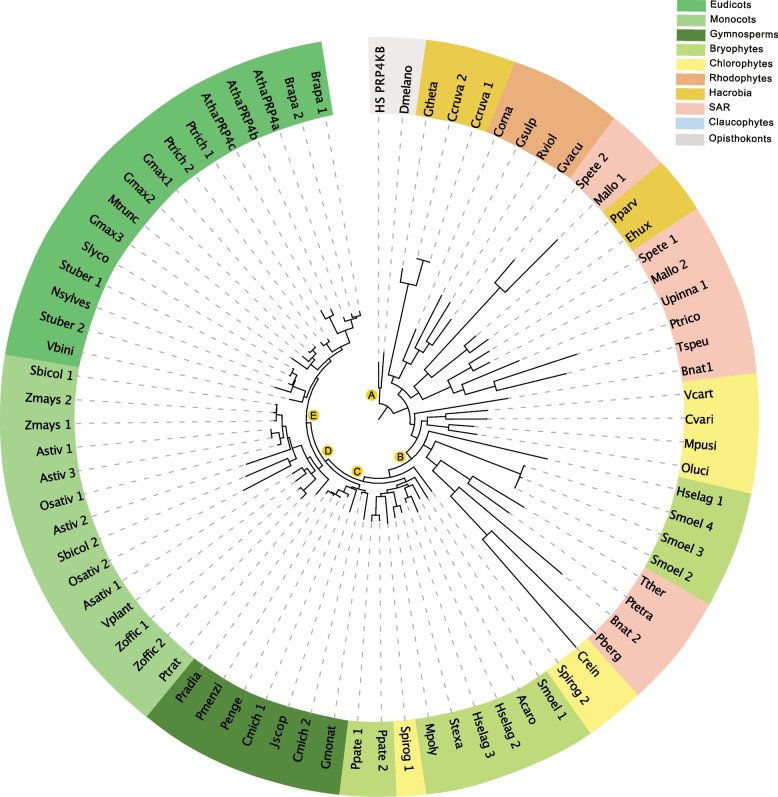
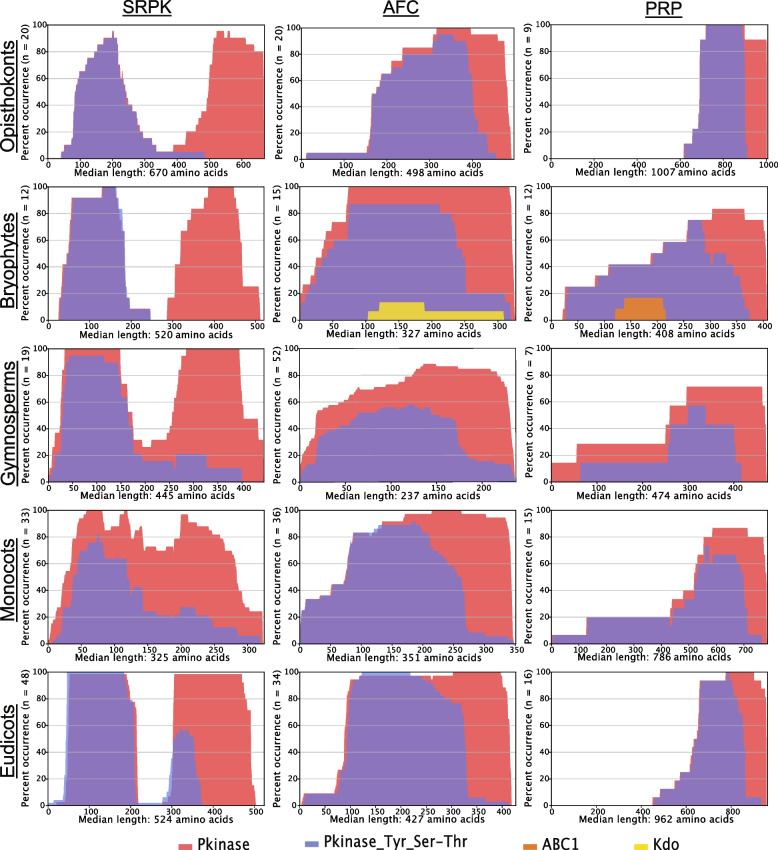
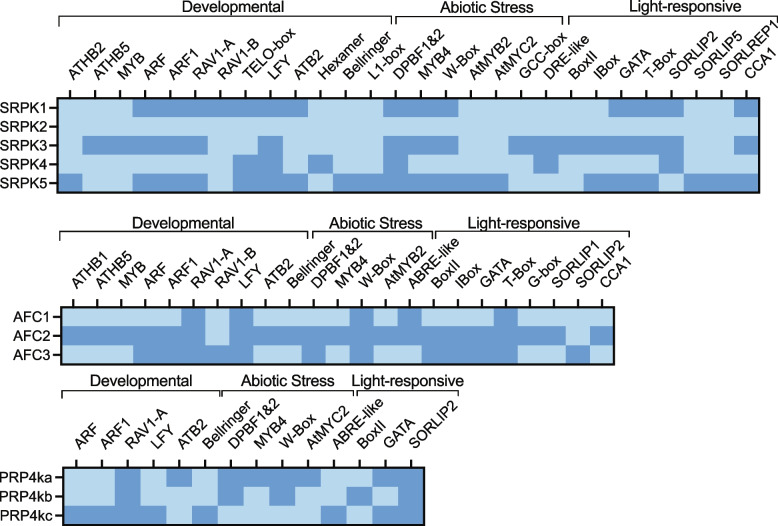
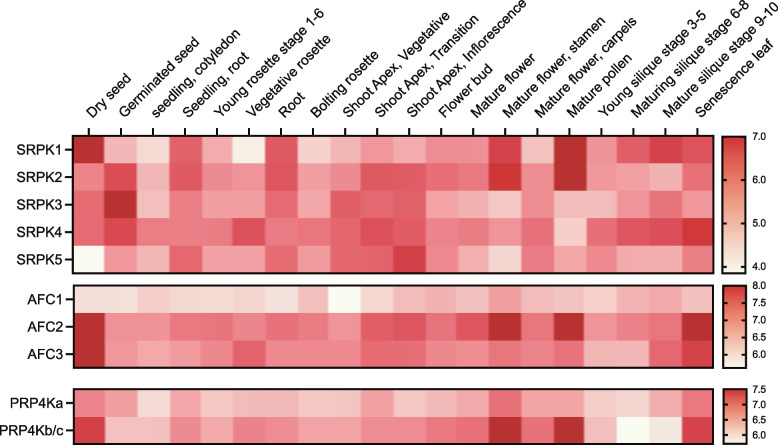
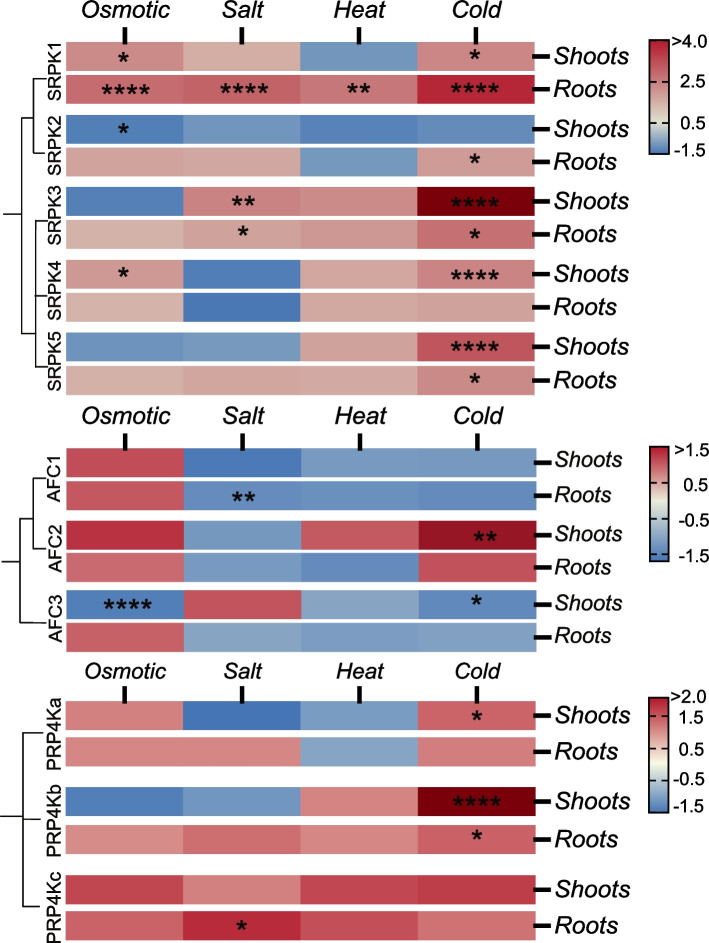
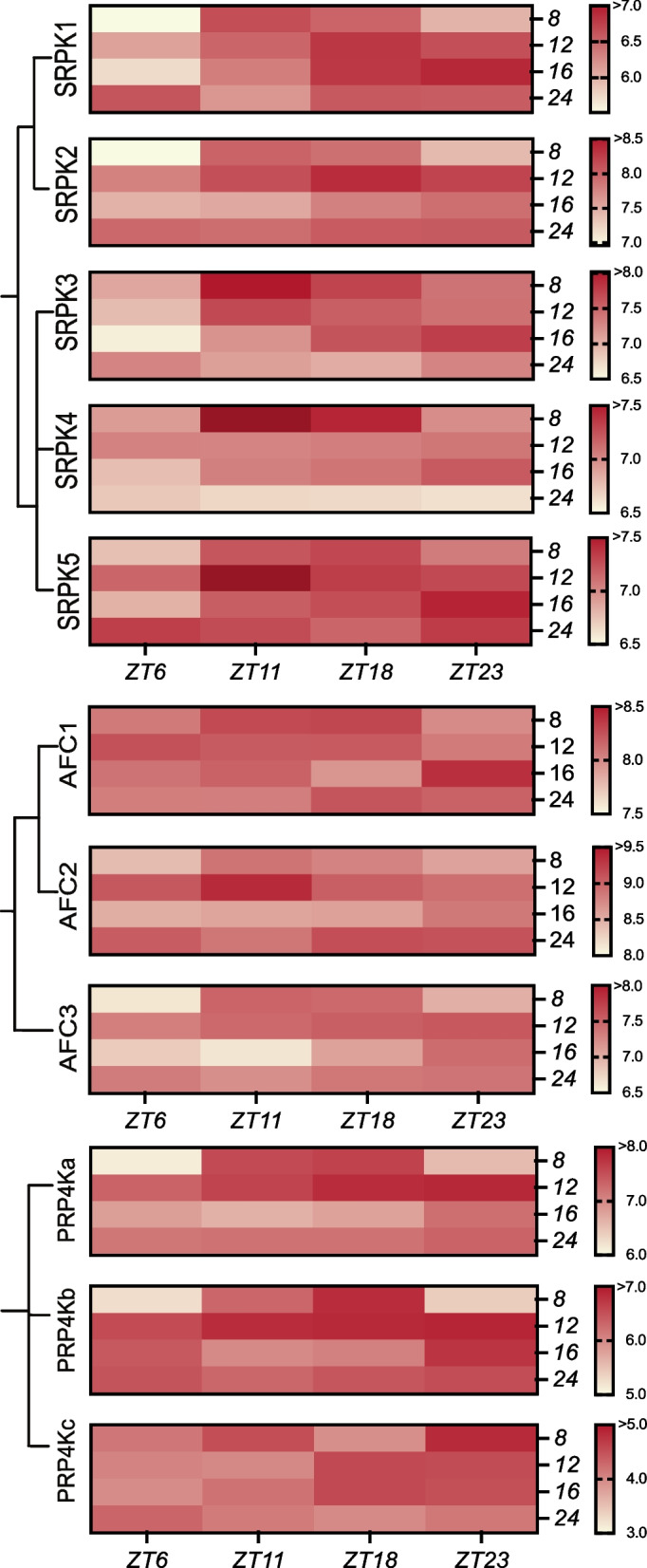
Nearly 60 - 80 % of intron-containing plant genes undergo alternative splicing in response to either stress or plant developmental cues. RNA splicing is performed by a large ribonucleoprotein complex called the spliceosome in conjunction with associated subunits such as serine arginine (SR) proteins, all of which undergo extensive phosphorylation. In plants, there are three main protein kinase families suggested to phosphorylate core spliceosome subunits and related splicing factors based on orthology to human splicing-related kinases: the SERINE/ARGININE PROTEIN KINASES (SRPK), ARABIDOPSIS FUS3 COMPLEMENT (AFC), and Pre-mRNA PROCESSING FACTOR 4 (PRP4K) protein kinases. To better define the conservation and role(s) of these kinases in plants, we performed a genome-scale analysis of the three families across photosynthetic eukaryotes, followed by extensive transcriptomic and bioinformatic analysis of all Arabidopsis thaliana SRPK, AFC, and PRP4K protein kinases to elucidate their biological functions. Unexpectedly, this revealed the existence of SRPK and AFC phylogenetic groups with distinct promoter elements and patterns of transcriptional response to abiotic stress, while PRP4Ks possess no phylogenetic sub-divisions, suggestive of functional redundancy. We also reveal splicing-related kinase families are both diel and photoperiod regulated, implicating different orthologs as discrete time-of-day RNA splicing regulators. This foundational work establishes a number of new hypotheses regarding how reversible spliceosome phosphorylation contributes to both diel plant cell regulation and abiotic stress adaptation in plants.
Keywords: Abiotic Stress; Diel cycle; Evolution; Protein kinases; mRNA splicing.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
Similar articles
-
Arabidopsis SME1 Regulates Plant Development and Response to Abiotic Stress by Determining Spliceosome Activity Specificity.Plant Cell. 2019 Feb;31(2):537-554. doi: 10.1105/tpc.18.00689. Epub 2019 Jan 29. Plant Cell. 2019. PMID: 30696706 Free PMC article.
-
Pre-mRNA splicing repression triggers abiotic stress signaling in plants.Plant J. 2017 Jan;89(2):291-309. doi: 10.1111/tpj.13383. Epub 2017 Jan 17. Plant J. 2017. PMID: 27664942
-
Genome-Wide Identification of Cassava Serine/Arginine-Rich Proteins: Insights into Alternative Splicing of Pre-mRNAs and Response to Abiotic Stress.Plant Cell Physiol. 2020 Jan 1;61(1):178-191. doi: 10.1093/pcp/pcz190. Plant Cell Physiol. 2020. PMID: 31596482
-
Alternative splicing in plant abiotic stress responses.Biochem Soc Trans. 2020 Oct 30;48(5):2117-2126. doi: 10.1042/BST20200281. Biochem Soc Trans. 2020. PMID: 32869832 Review.
-
Plant serine/arginine-rich proteins: roles in precursor messenger RNA splicing, plant development, and stress responses.Wiley Interdiscip Rev RNA. 2011 Nov-Dec;2(6):875-89. doi: 10.1002/wrna.98. Epub 2011 Jul 15. Wiley Interdiscip Rev RNA. 2011. PMID: 21766458 Review.
Cited by
-
Importance of pre-mRNA splicing and its study tools in plants.Adv Biotechnol (Singap). 2024 Feb 8;2(1):4. doi: 10.1007/s44307-024-00009-9. Adv Biotechnol (Singap). 2024. PMID: 39883322 Free PMC article. Review.
-
C-terminus of serine-arginine protein kinase-like protein, SrpkF, is involved in conidiophore formation and hyphal growth under salt stress in Aspergillus aculeatus.Int Microbiol. 2024 Feb;27(1):91-100. doi: 10.1007/s10123-023-00373-x. Epub 2023 May 17. Int Microbiol. 2024. PMID: 37195349
-
Phosphorylation mediated regulation of RNA splicing in plants.Front Plant Sci. 2023 Sep 14;14:1249057. doi: 10.3389/fpls.2023.1249057. eCollection 2023. Front Plant Sci. 2023. PMID: 37780493 Free PMC article. Review.
-
Correction: Genome-scale analysis of Arabidopsis splicing-related protein kinase families reveals roles in abiotic stress adaptation.BMC Plant Biol. 2023 May 23;23(1):272. doi: 10.1186/s12870-023-04289-6. BMC Plant Biol. 2023. PMID: 37217886 Free PMC article. No abstract available.
-
A cell fractionation and quantitative proteomics pipeline to enable functional analyses of cotton fiber development.Plant J. 2025 Feb;121(4):e17246. doi: 10.1111/tpj.17246. Plant J. 2025. PMID: 39970036 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
