

Scaffold proteins as dynamic integrators of biological processes
- PMID: 36273588
- PMCID: PMC9672449
- DOI: 10.1016/j.jbc.2022.102628
Scaffold proteins as dynamic integrators of biological processes
Abstract
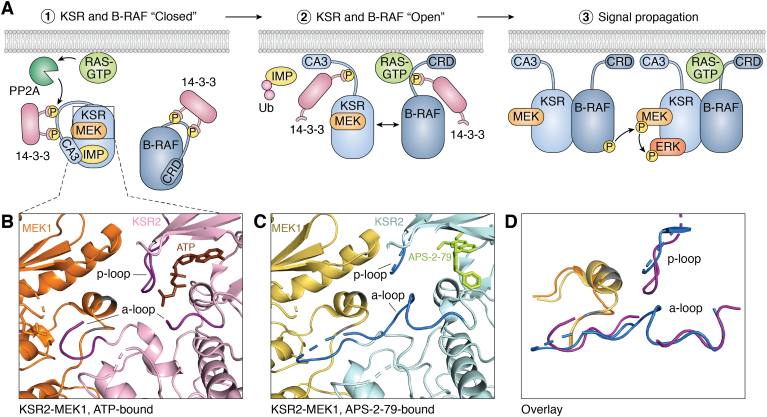
Scaffold proteins act as molecular hubs for the docking of multiple proteins to organize efficient functional units for signaling cascades. Over 300 human proteins have been characterized as scaffolds, acting in a variety of signaling pathways. While the term scaffold implies a static, supportive platform, it is now clear that scaffolds are not simply inert docking stations but can undergo conformational changes that affect their dependent signaling pathways. In this review, we catalog scaffold proteins that have been shown to undergo actionable conformational changes, with a focus on the role that conformational change plays in the activity of the classic yeast scaffold STE5, as well as three human scaffold proteins (KSR, NEMO, SHANK3) that are integral to well-known signaling pathways (RAS, NF-κB, postsynaptic density). We also discuss scaffold protein conformational changes vis-à-vis liquid-liquid phase separation. Changes in scaffold structure have also been implicated in human disease, and we discuss how aberrant conformational changes may be involved in disease-related dysregulation of scaffold and signaling functions. Finally, we discuss how understanding these conformational dynamics will provide insight into the flexibility of signaling cascades and may enhance our ability to treat scaffold-associated diseases.
Keywords: KSR; NEMO; SHANK3; STE5; conformational change; liquid–liquid phase separation; scaffold protein; signaling.
Copyright © 2022 The Authors. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Conflict of interest The authors declare that they have no conflicts of interest with the contents of this article.
Figures



References
-
- Shaw A.S., Filbert E.L. Scaffold proteins and immune-cell signalling. Nat. Rev. Immunol. 2009;9:47–56. - PubMed
-
- Buday L., Tompa P. Functional classification of scaffold proteins and related molecules. FEBS J. 2010;277:4348–4355. - PubMed
-
- Alexa A., Varga J., Reményi A. Scaffolds are 'active' regulators of signaling modules. FEBS J. 2010;277:4376–4382. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
