

GLH-1/Vasa represses neuropeptide expression and drives spermiogenesis in the C. elegans germline
- PMID: 36273621
- PMCID: PMC9677334
- DOI: 10.1016/j.ydbio.2022.10.003
GLH-1/Vasa represses neuropeptide expression and drives spermiogenesis in the C. elegans germline
Abstract
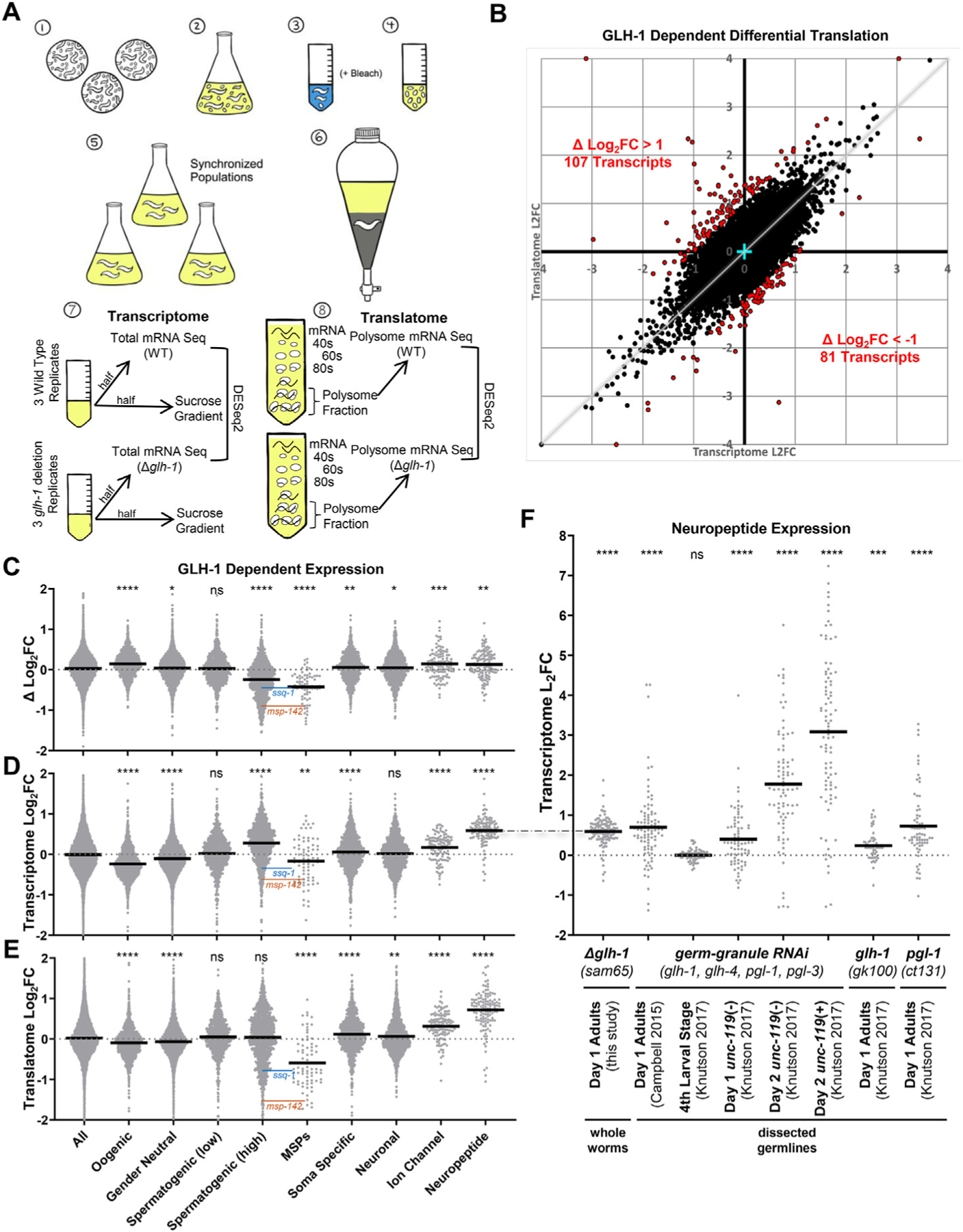
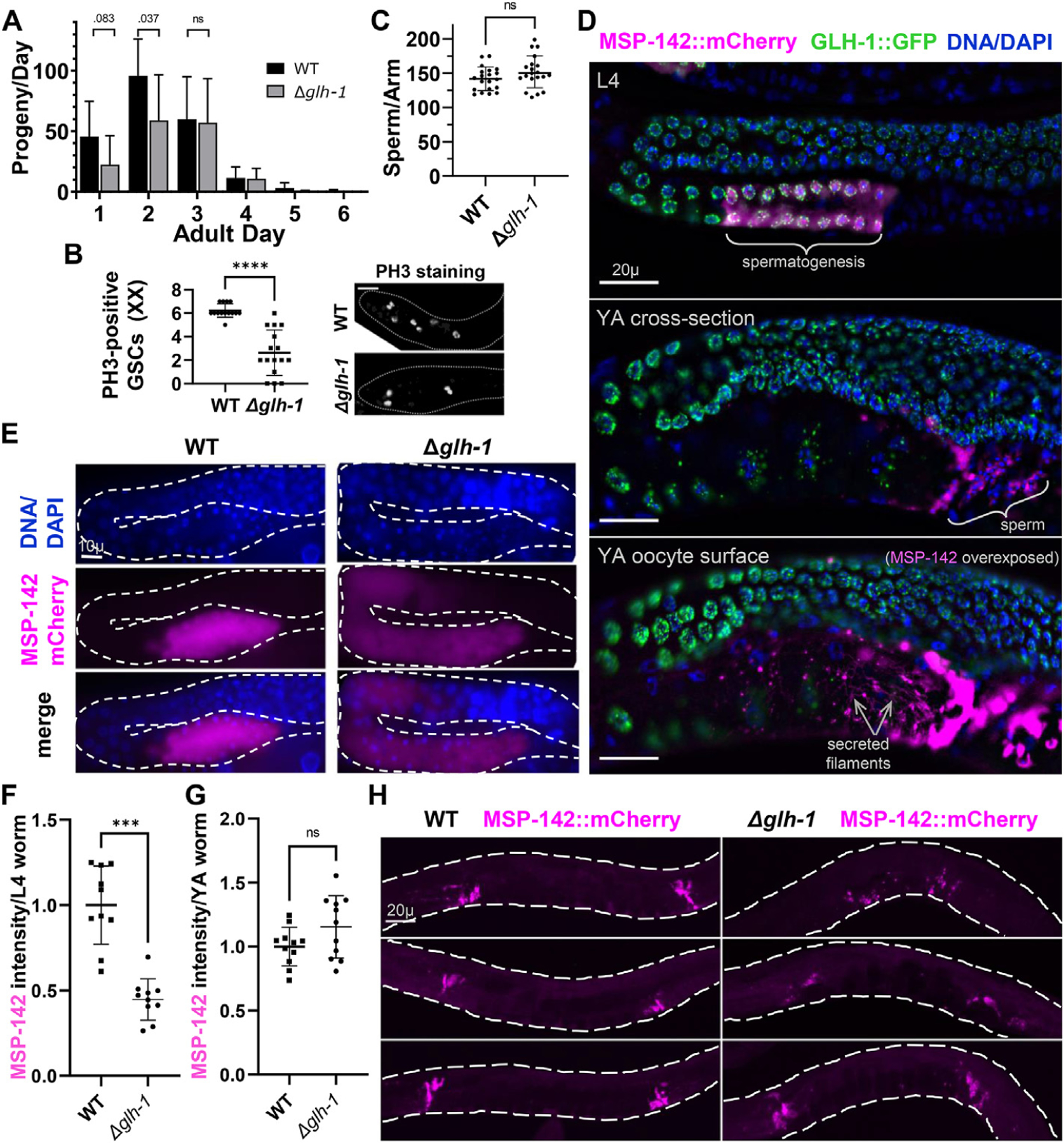
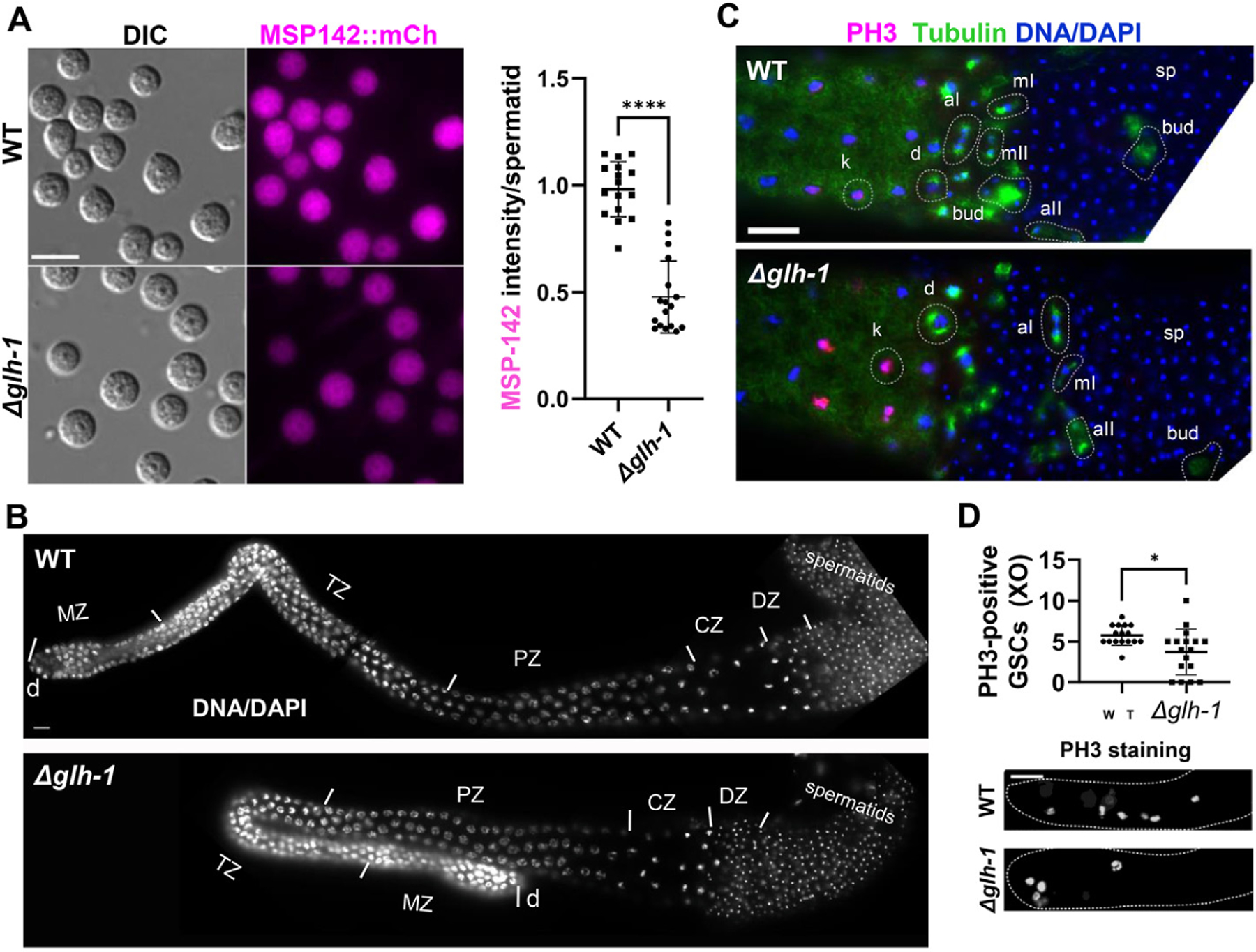
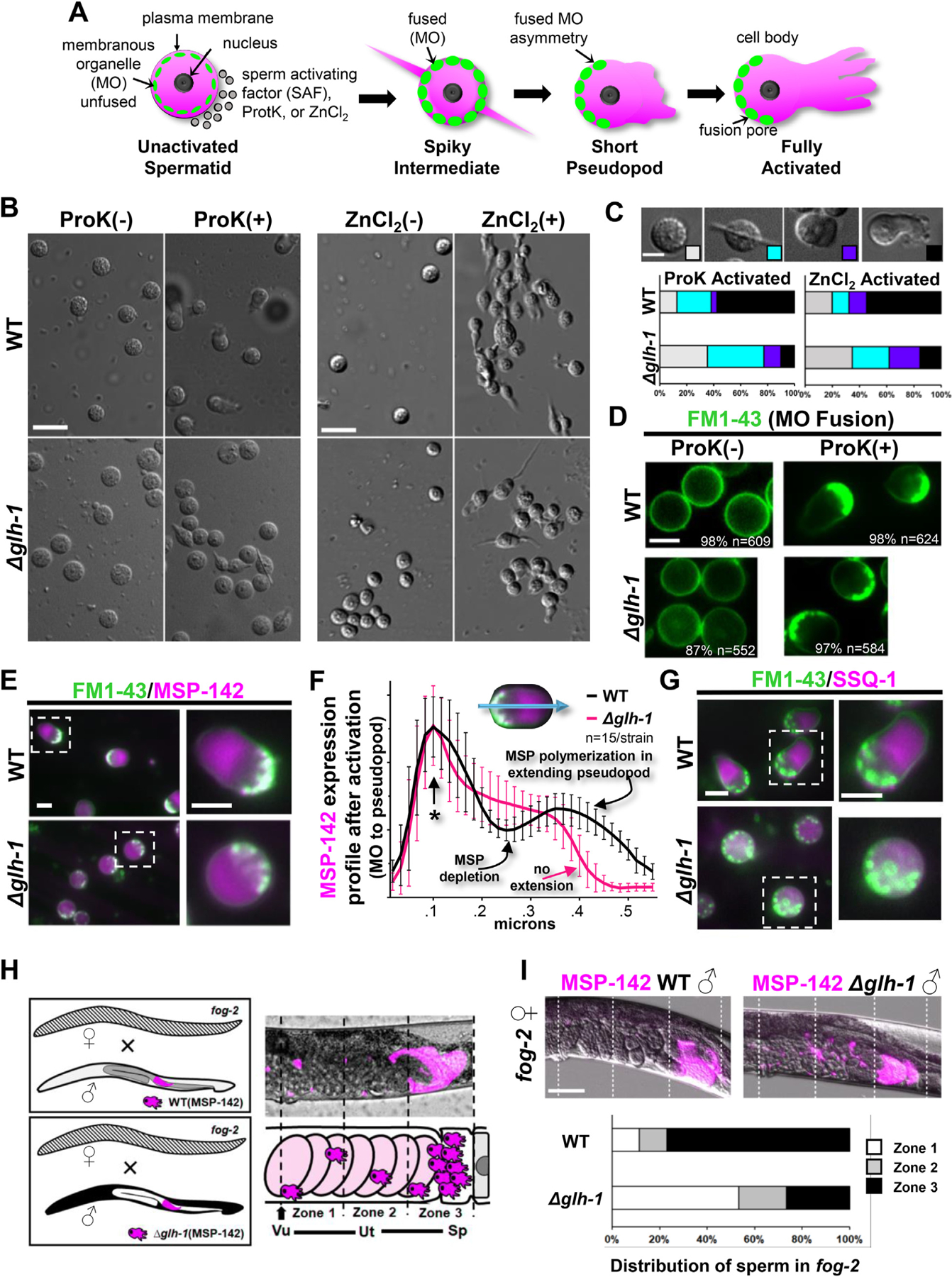
Germ granules harbor processes that maintain germline integrity and germline stem cell capacity. Depleting core germ granule components in C. elegans leads to the reprogramming of germ cells, causing them to express markers of somatic differentiation in day-two adults. Somatic reprogramming is associated with complete sterility at this stage. The resulting germ cell atrophy and other pleiotropic defects complicate our understanding of the initiation of reprogramming and how processes within germ granules safeguard the totipotency and immortal potential of germline stem cells. To better understand the initial events of somatic reprogramming, we examined total mRNA (transcriptome) and polysome-associated mRNA (translatome) changes in a precision full-length deletion of glh-1, which encodes a homolog of the germline-specific Vasa/DDX4 DEAD-box RNA helicase. Fertile animals at a permissive temperature were analyzed as young adults, a stage that precedes by 24 h the previously determined onset of somatic reporter-gene expression in the germline. Two significant changes are observed at this early stage. First, the majority of neuropeptide-encoding transcripts increase in both the total and polysomal mRNA fractions, suggesting that GLH-1 or its effectors suppress this expression. Second, there is a significant decrease in Major Sperm Protein (MSP)-domain mRNAs when glh-1 is deleted. We find that the presence of GLH-1 helps repress spermatogenic expression during oogenesis, but boosts MSP expression to drive spermiogenesis and sperm motility. These insights define an early role for GLH-1 in repressing somatic reprogramming to maintain germline integrity.
Keywords: C. elegans; DDX4; GLH-1; Germ granules; Germline; MSP; Major sperm protein; Neuropeptides; P granules; Polysomes; Sperm; Spermiogenesis; Vasa.
Copyright © 2022 Elsevier Inc. All rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
