

A computational solution for bolstering reliability of epigenetic clocks: Implications for clinical trials and longitudinal tracking
- PMID: 36277076
- PMCID: PMC9586209
- DOI: 10.1038/s43587-022-00248-2
A computational solution for bolstering reliability of epigenetic clocks: Implications for clinical trials and longitudinal tracking
Abstract
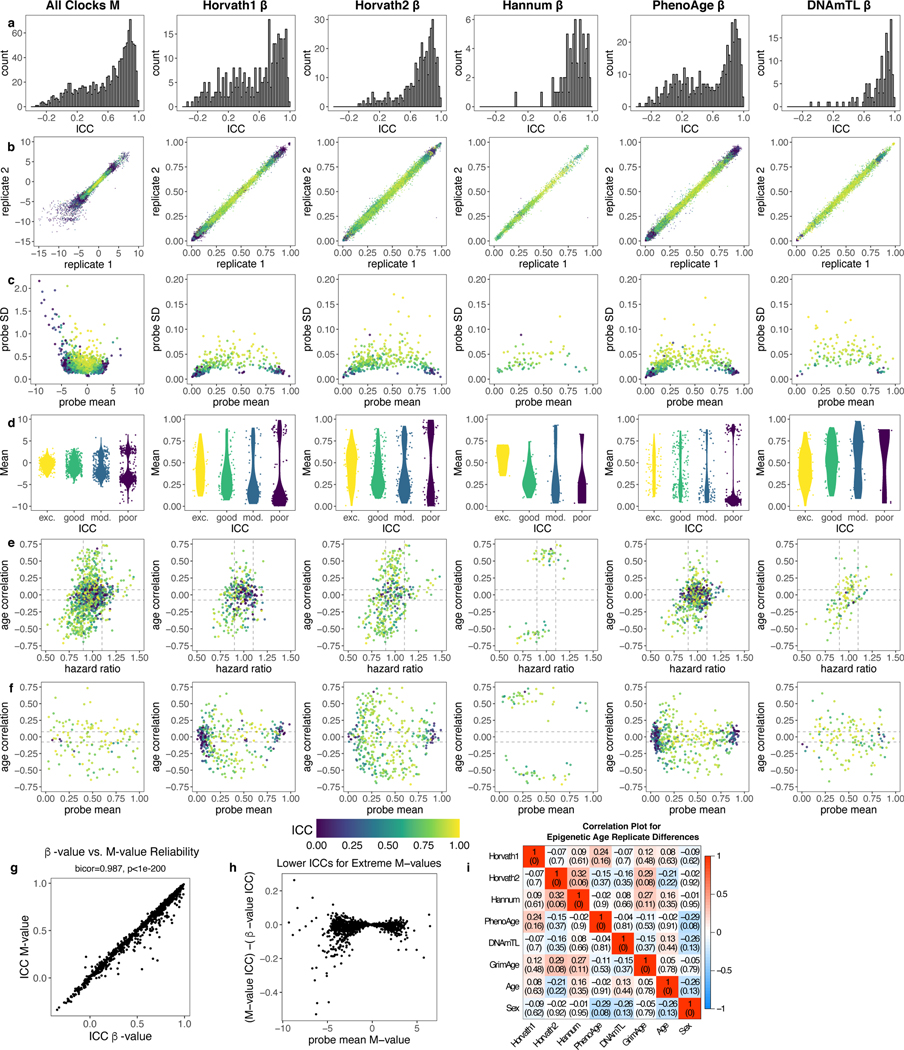
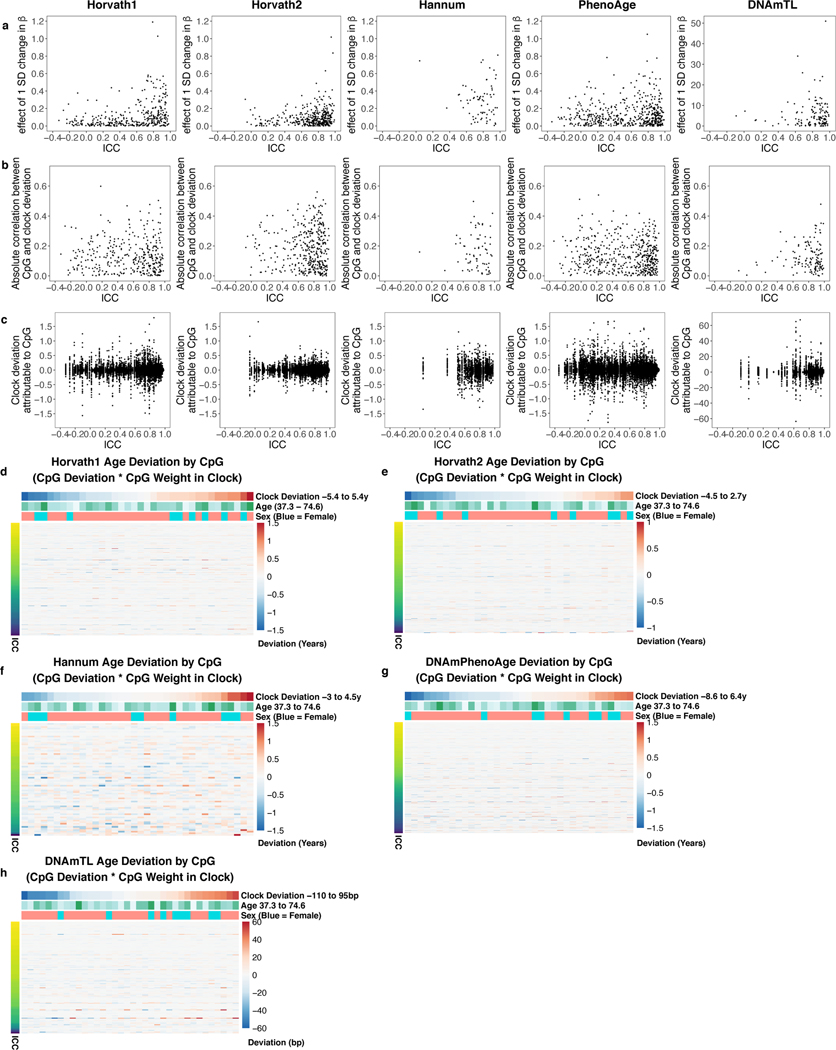
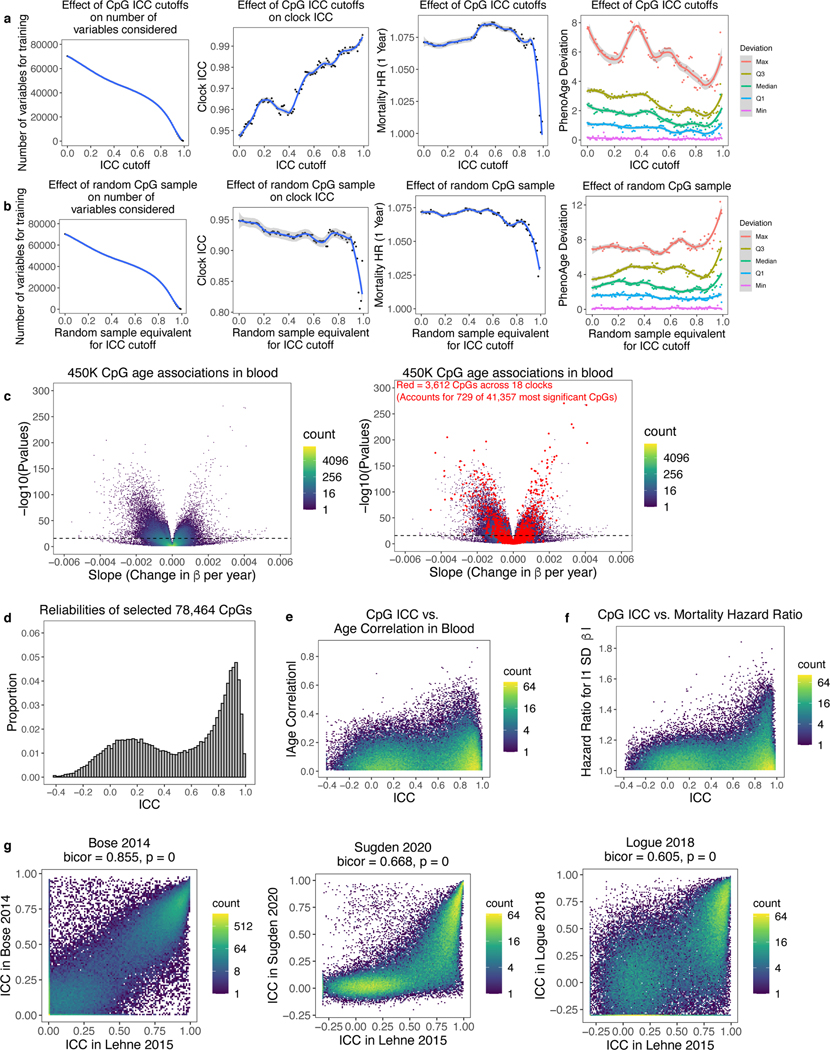
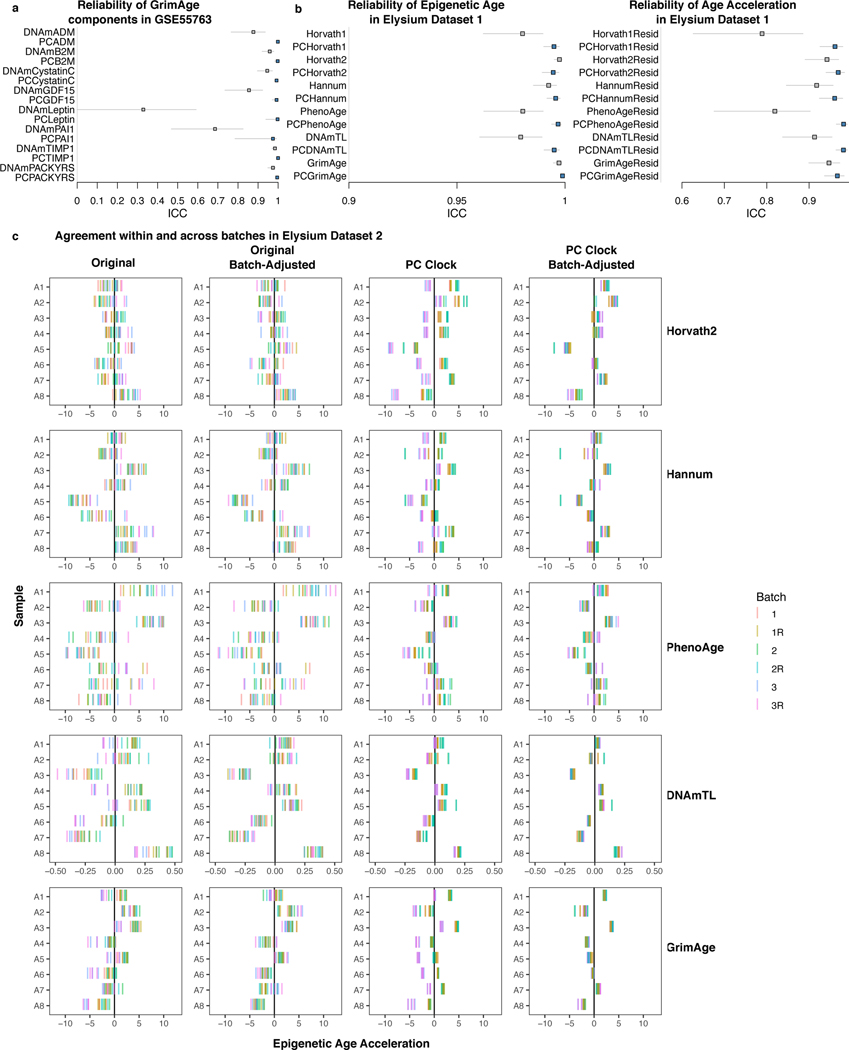
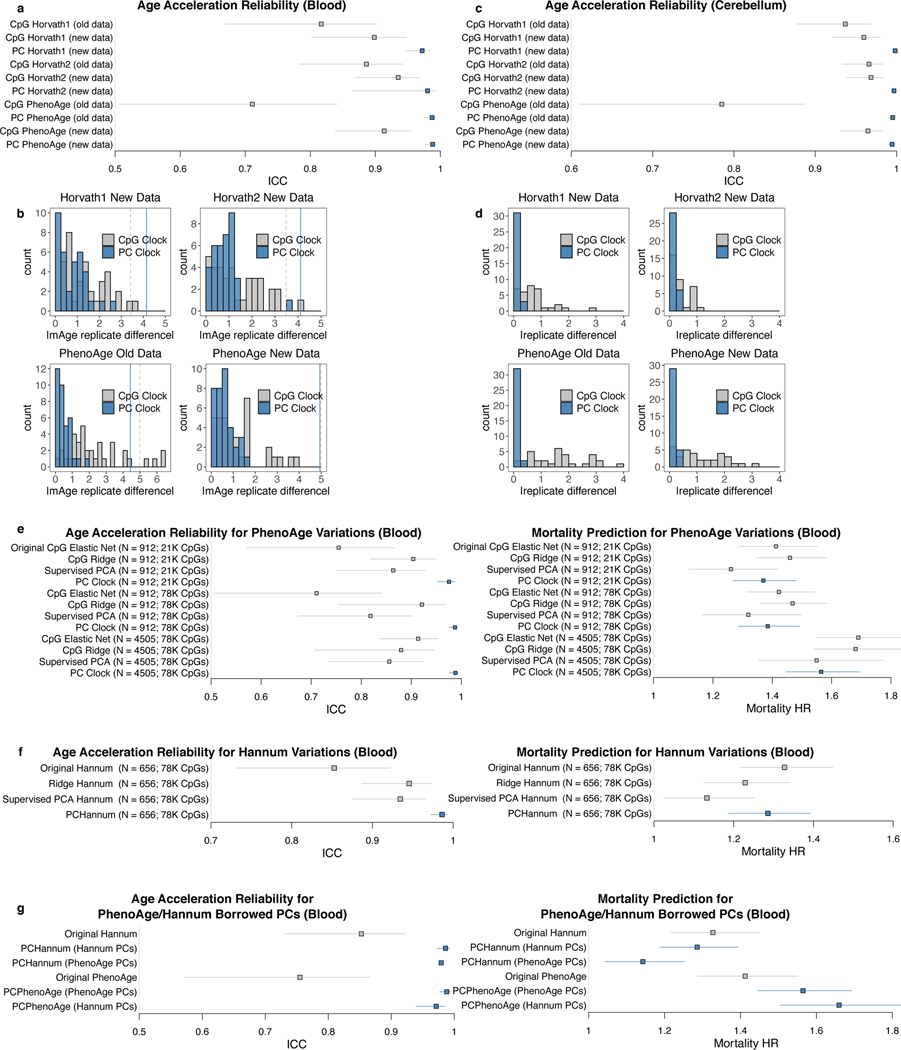
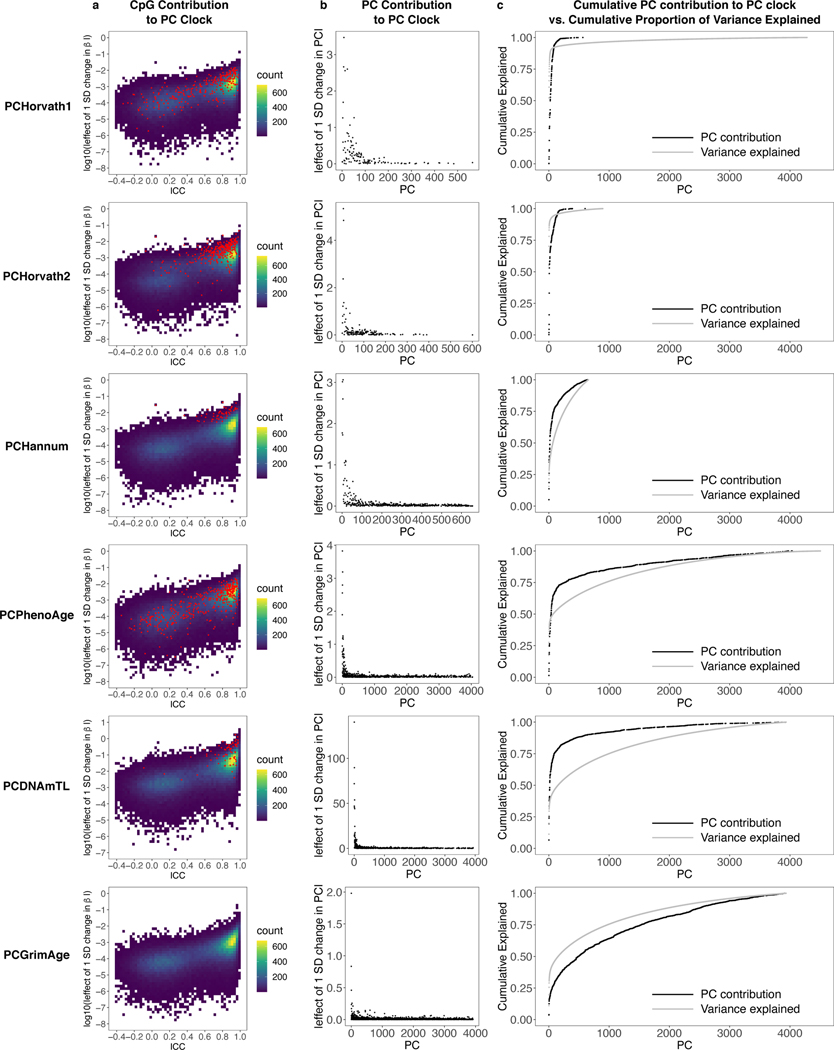
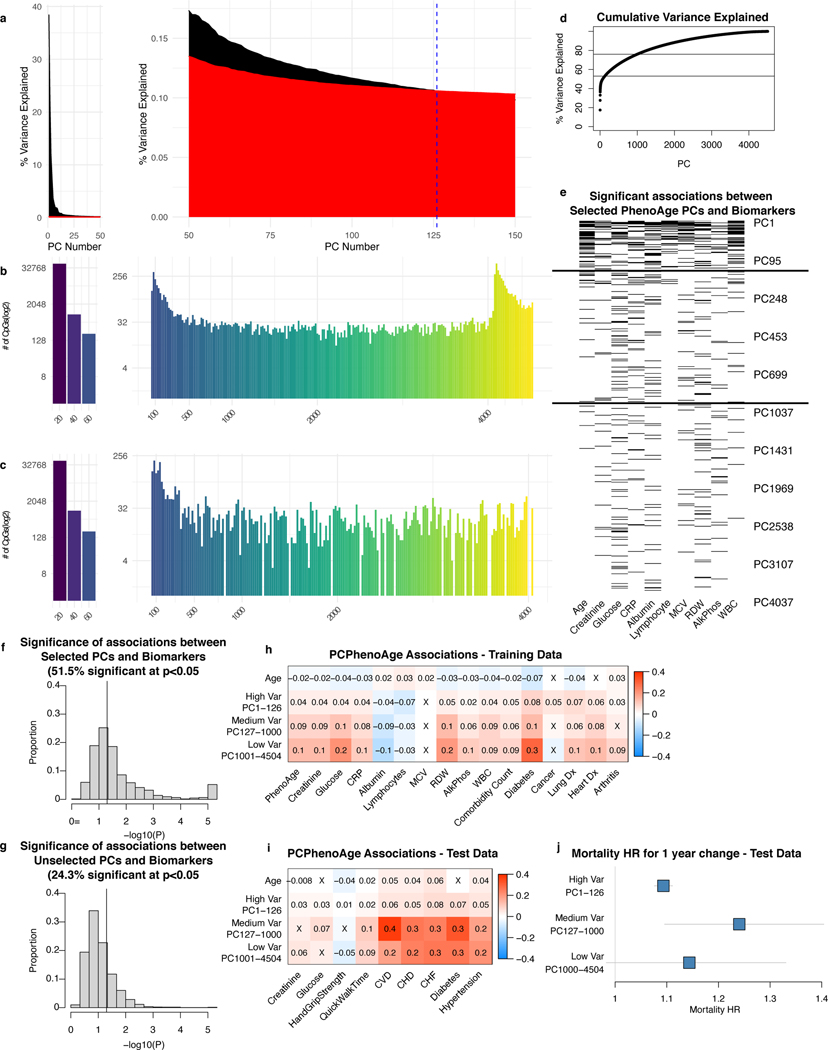
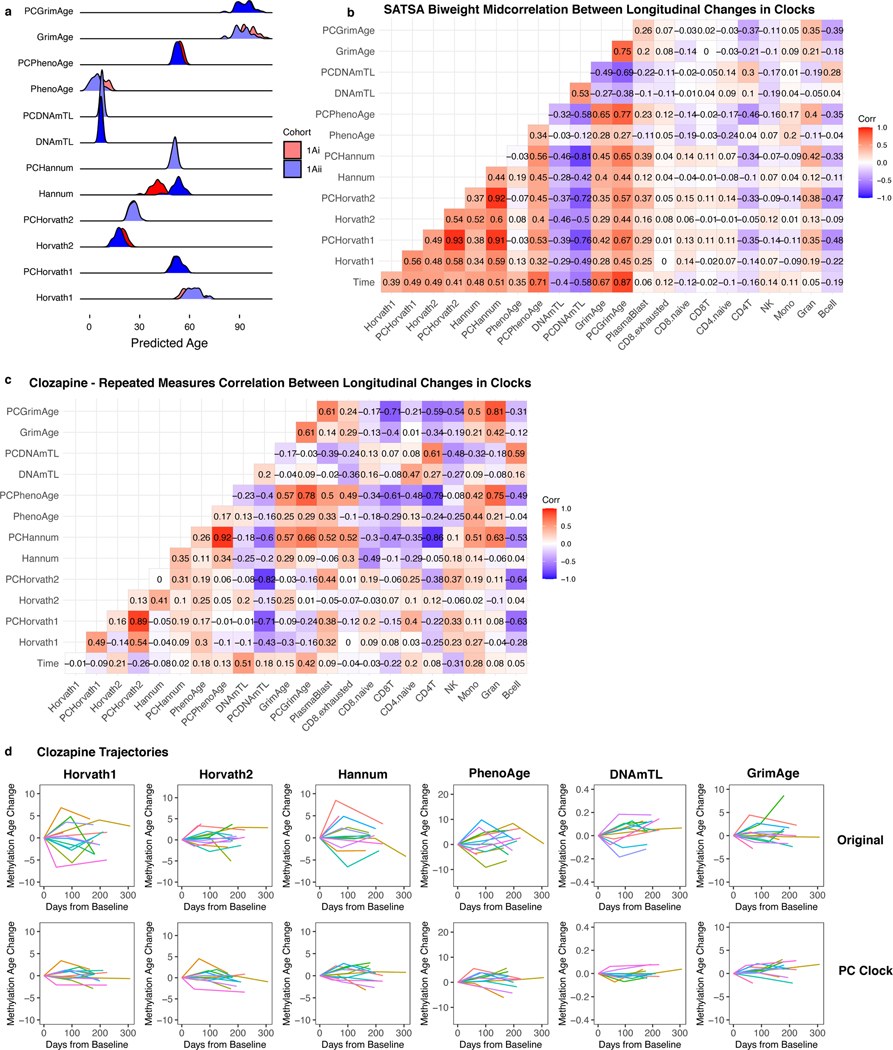
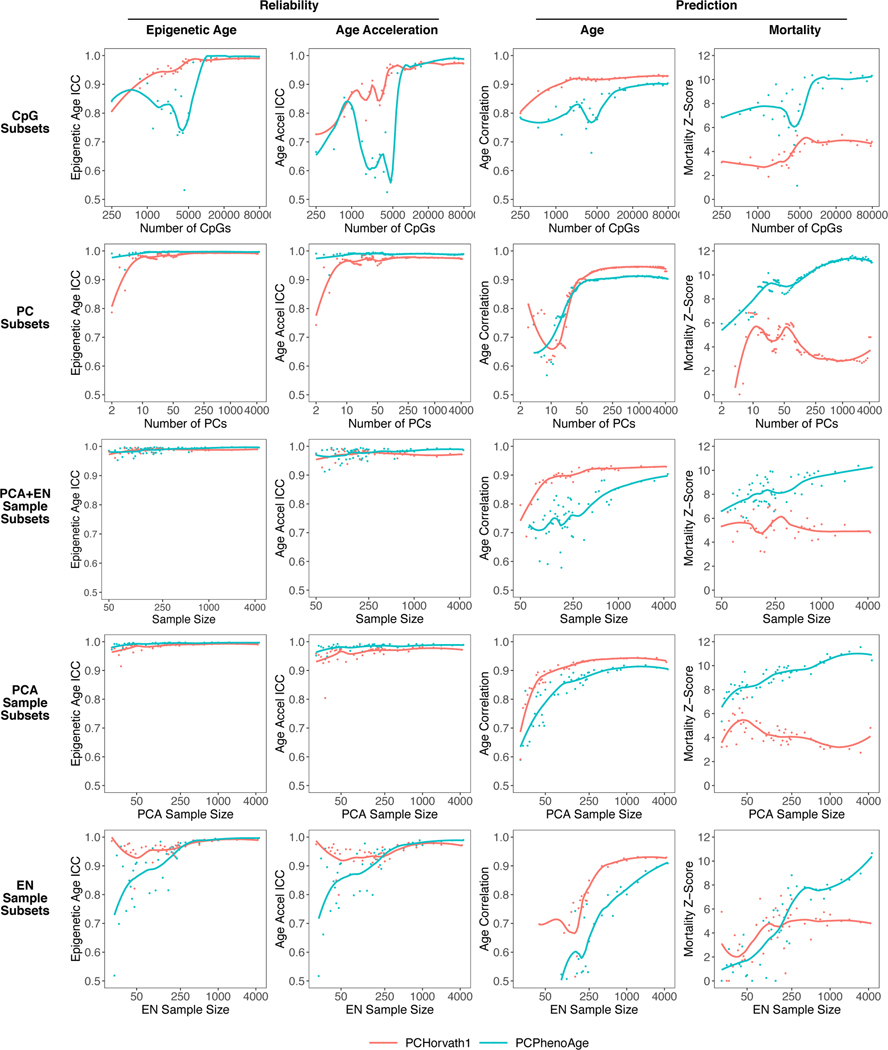
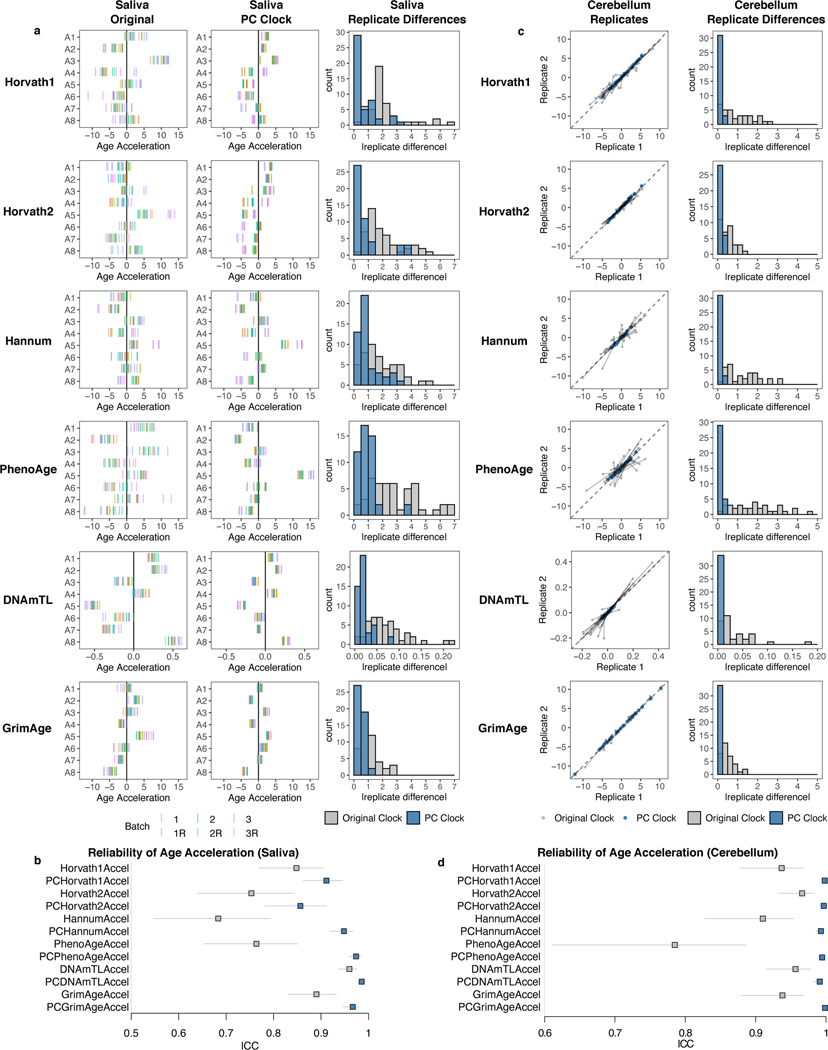
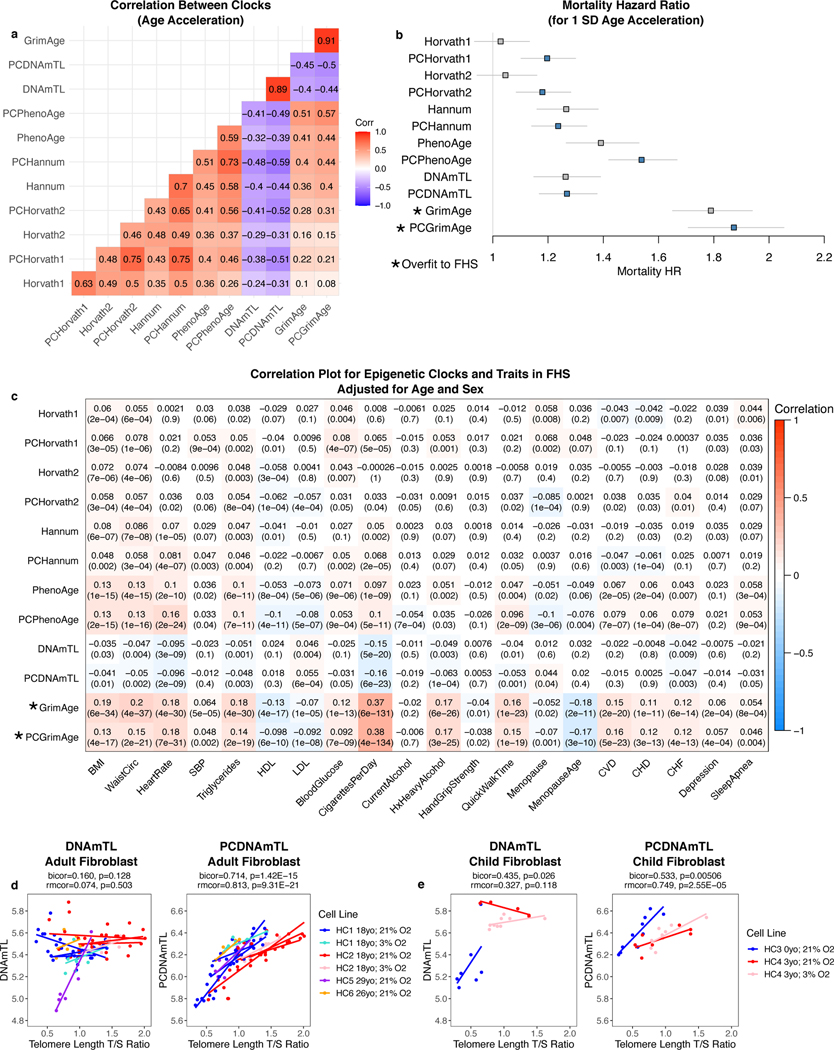
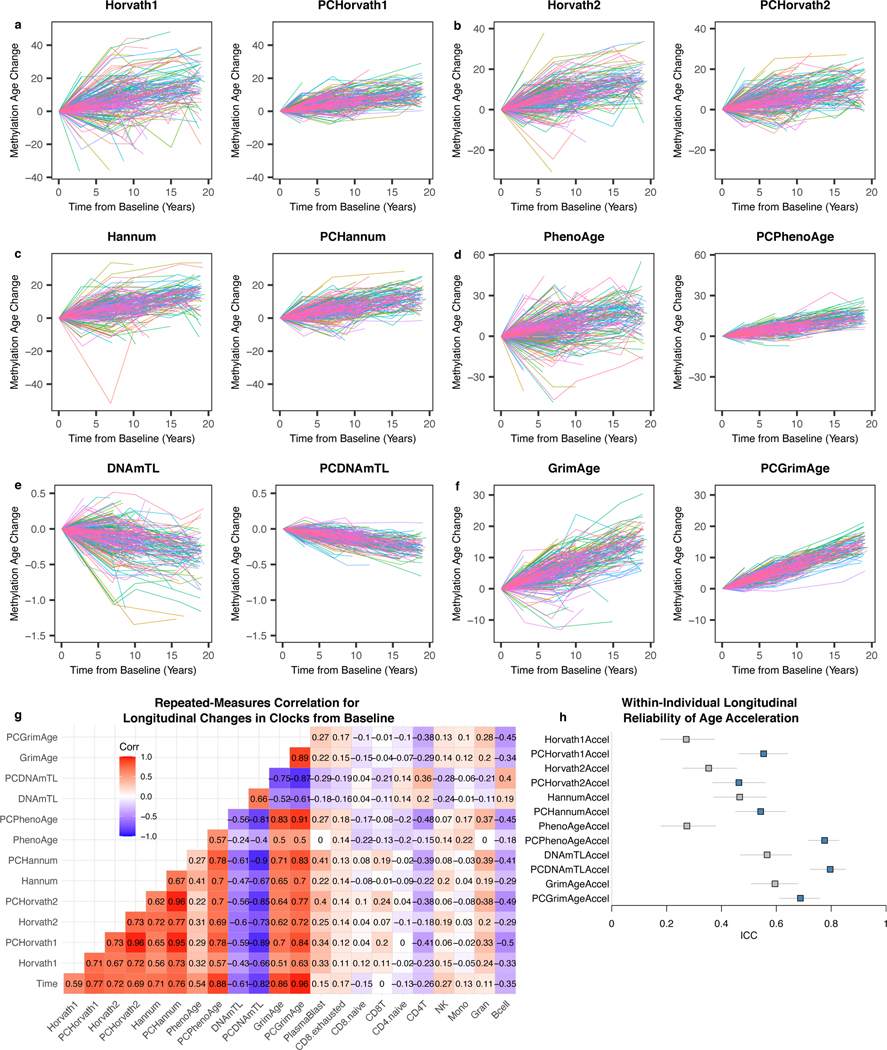
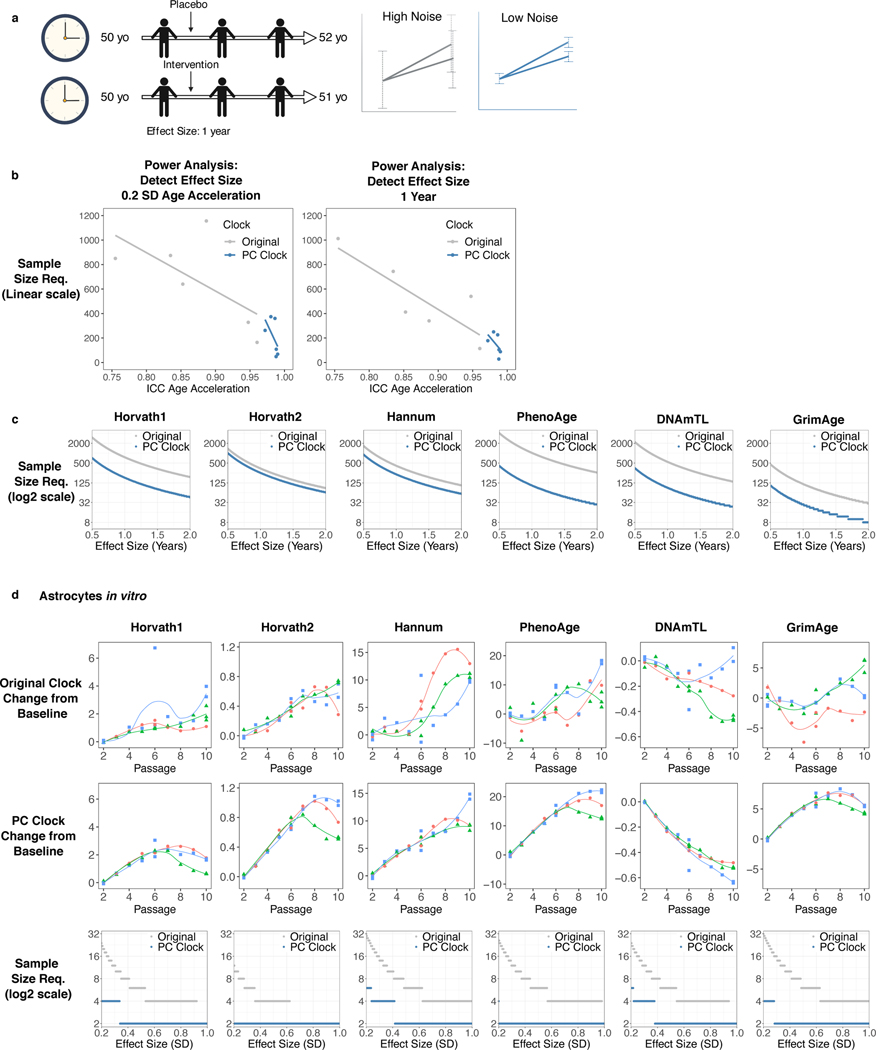
Epigenetic clocks are widely used aging biomarkers calculated from DNA methylation data, but this data can be surprisingly unreliable. Here we show technical noise produces deviations up to 9 years between replicates for six prominent epigenetic clocks, limiting their utility. We present a computational solution to bolster reliability, calculating principal components from CpG-level data as input for biological age prediction. Our retrained principal-component versions of six clocks show agreement between most replicates within 1.5 years, improved detection of clock associations and intervention effects, and reliable longitudinal trajectories in vivo and in vitro. This method entails only one additional step compared to traditional clocks, requires no replicates or prior knowledge of CpG reliabilities for training, and can be applied to any existing or future epigenetic biomarker. The high reliability of principal component-based clocks is critical for applications to personalized medicine, longitudinal tracking, in vitro studies, and clinical trials of aging interventions.
Keywords: aging; biomarker; epigenetic clock; longitudinal analysis; reliability.
Conflict of interest statement
Competing Interests Statement MEL and AHC have built epigenetic aging metrics involving the technology described in the present manuscript, and these metrics are licensed by Elysium Health through Yale University. Elysium provided paired blood and saliva replicate datasets reported in this study, but otherwise did not fund the study and did not play a role in conceptualization, design, decision to publish, or preparation of the manuscript. MEL previously acted as a Scientific Advisor for, and received consulting fees from, Elysium Health, Inc. THS was previously an employee of Elysium Health, Inc. AHC received consulting fees from FOXO Technologies, Inc. for work unrelated to the present manuscript. All other authors report no biomedical financial interests or potential conflicts of interest.
Figures
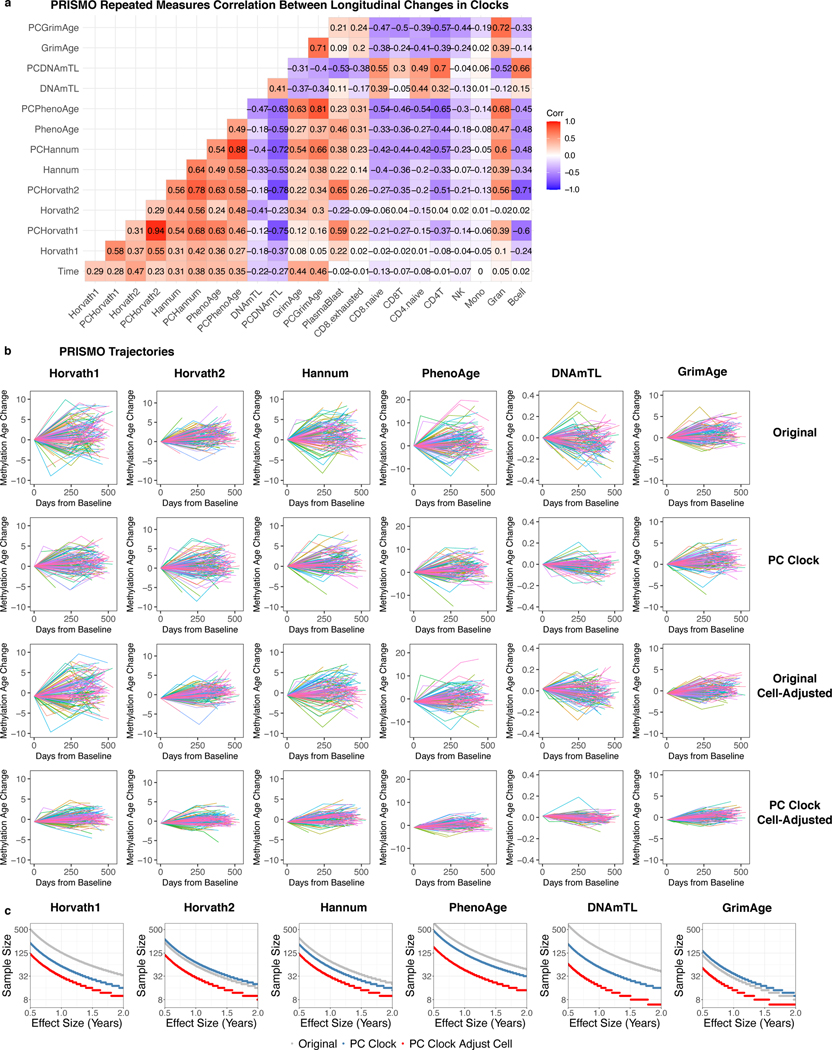
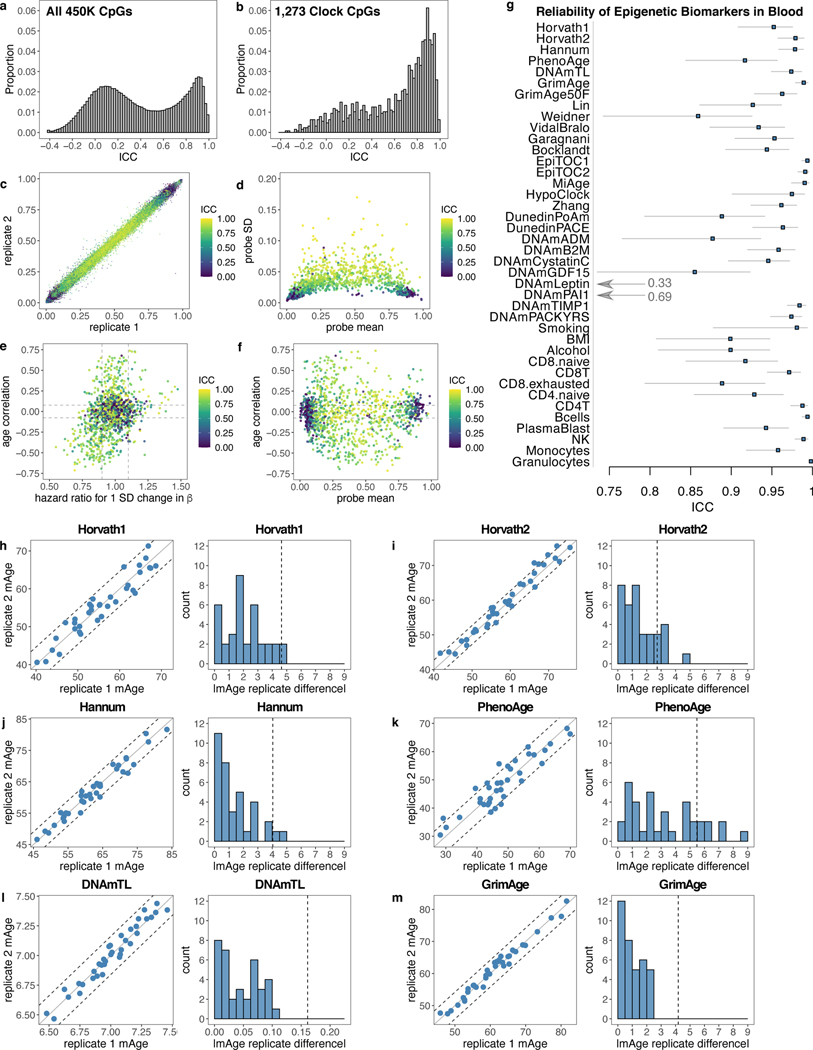
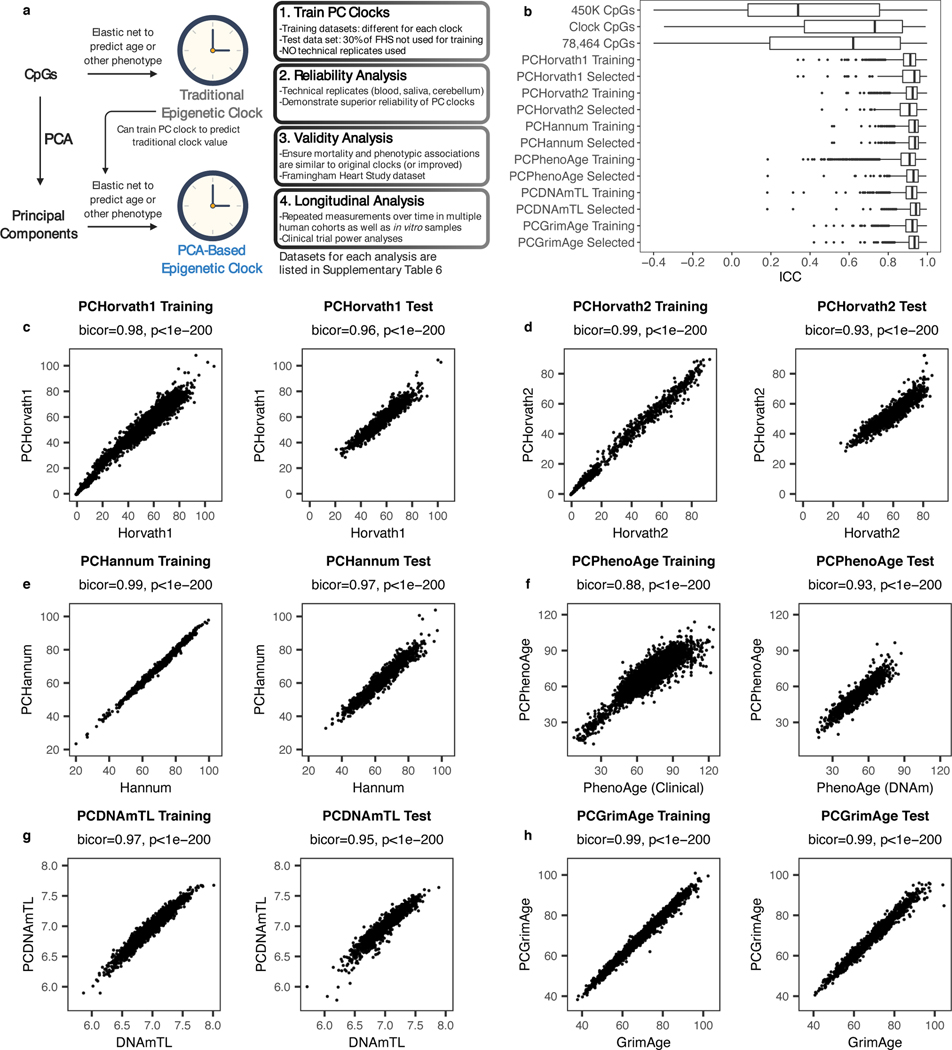
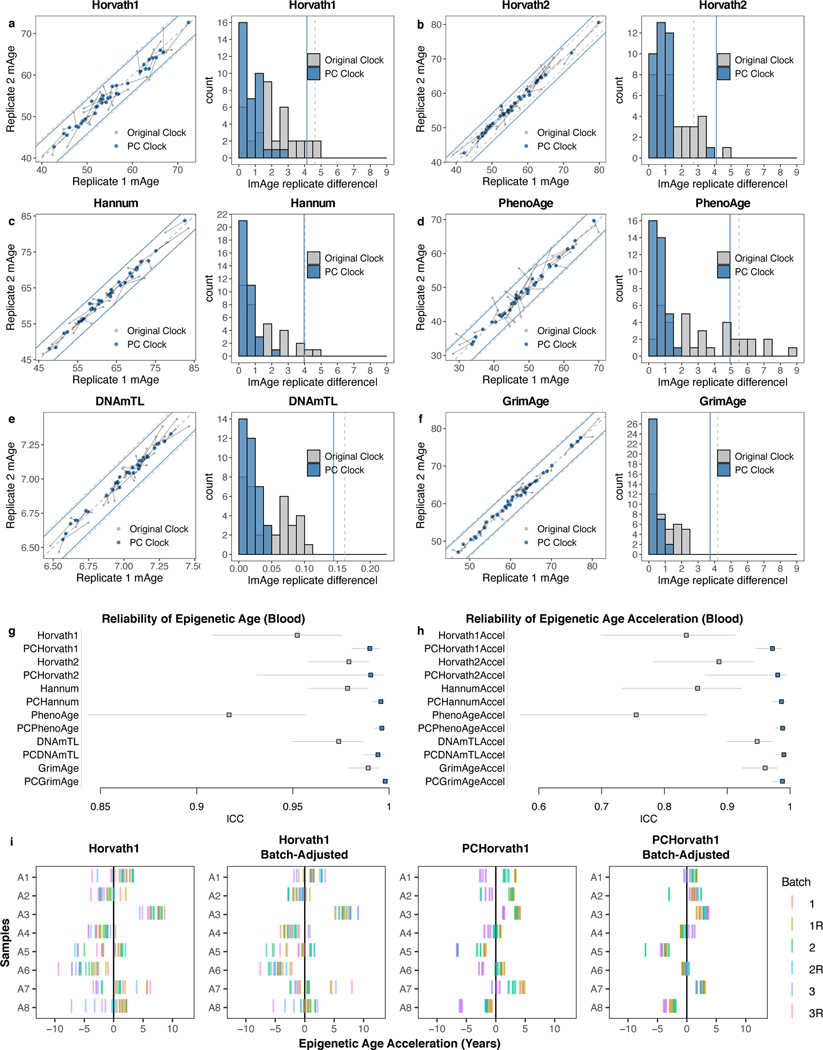
References
-
- Horvath S. & Raj K. DNA methylation-based biomarkers and the epigenetic clock theory of ageing. Nat. Rev. Genet. 19, 371–384 (2018). - PubMed
Publication types
MeSH terms
Grants and funding
- R01 AG068285/AG/NIA NIH HHS/United States
- R01 AG066828/AG/NIA NIH HHS/United States
- R01 AG060110/AG/NIA NIH HHS/United States
- R01 MD009164/MD/NIMHD NIH HHS/United States
- P30 AG066508/AG/NIA NIH HHS/United States
- T32 MH019961/MH/NIMH NIH HHS/United States
- R01 AG028555/AG/NIA NIH HHS/United States
- N01AG50002/DA/NIDA NIH HHS/United States
- P30 AG017265/AG/NIA NIH HHS/United States
- R01 AG057912/AG/NIA NIH HHS/United States
- U01 AG009740/AG/NIA NIH HHS/United States
- R01 AG065403/AG/NIA NIH HHS/United States
- R01 AG010175/AG/NIA NIH HHS/United States
- R01 AG068937/AG/NIA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
