

Structural and dynamical determinants of a β-sheet-enriched intermediate involved in amyloid fibrillar assembly of human prion protein
- PMID: 36277622
- PMCID: PMC9473526
- DOI: 10.1039/d2sc00345g
Structural and dynamical determinants of a β-sheet-enriched intermediate involved in amyloid fibrillar assembly of human prion protein
Abstract
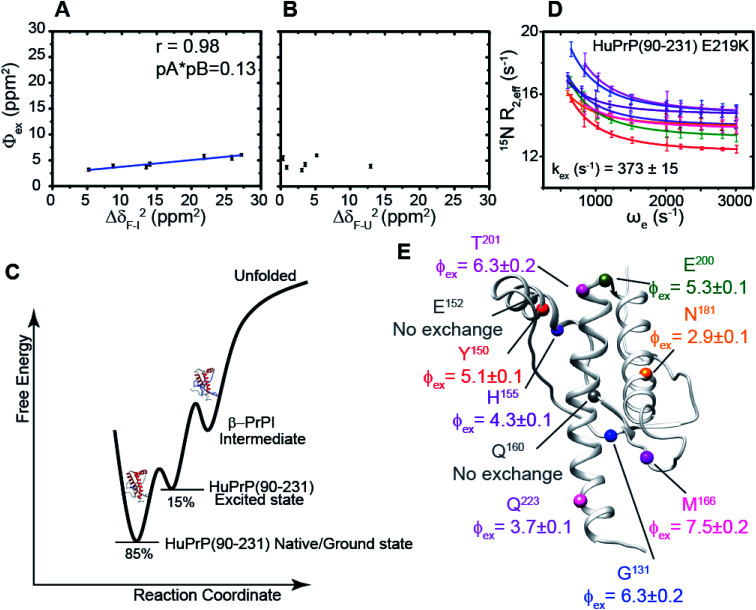
The conformational conversion of the cellular prion protein (PrPC) into a misfolded, aggregated and infectious scrapie isoform is associated with prion disease pathology and neurodegeneration. Despite the significant number of experimental and theoretical studies the molecular mechanism regulating this structural transition is still poorly understood. Here, via Nuclear Magnetic Resonance (NMR) methodologies we investigate at the atomic level the mechanism of the human HuPrP(90-231) thermal unfolding and characterize the conformational equilibrium between its native structure and a β-enriched intermediate state, named β-PrPI. By comparing the folding mechanisms of metal-free and Cu2+-bound HuPrP(23-231) and HuPrP(90-231) we show that the coupling between the N- and C-terminal domains, through transient electrostatic interactions, is the key molecular process in tuning long-range correlated μs-ms dynamics that in turn modulate the folding process. Moreover, via thioflavin T (ThT)-fluorescence fibrillization assays we show that β-PrPI is involved in the initial stages of PrP fibrillation, overall providing a clear molecular description of the initial phases of prion misfolding. Finally, we show by using Real-Time Quaking-Induced Conversion (RT-QuIC) that the β-PrPI acts as a seed for the formation of amyloid aggregates with a seeding activity comparable to that of human infectious prions.
This journal is © The Royal Society of Chemistry.
Conflict of interest statement
There are no conflicts to declare.
Figures






Similar articles
-
Unfolding Mechanism and Fibril Formation Propensity of Human Prion Protein in the Presence of Molecular Crowding Agents.Int J Mol Sci. 2024 Sep 13;25(18):9916. doi: 10.3390/ijms25189916. Int J Mol Sci. 2024. PMID: 39337404 Free PMC article.
-
Rapid end-point quantitation of prion seeding activity with sensitivity comparable to bioassays.PLoS Pathog. 2010 Dec 2;6(12):e1001217. doi: 10.1371/journal.ppat.1001217. PLoS Pathog. 2010. PMID: 21152012 Free PMC article.
-
Molecular insights into the critical role of gallate moiety of green tea catechins in modulating prion fibrillation, cellular internalization, and neuronal toxicity.Int J Biol Macromol. 2022 Dec 31;223(Pt A):755-765. doi: 10.1016/j.ijbiomac.2022.11.049. Epub 2022 Nov 9. Int J Biol Macromol. 2022. PMID: 36368361
-
NMR structural studies of human cellular prion proteins.Curr Top Med Chem. 2013;13(19):2407-18. doi: 10.2174/15680266113136660169. Curr Top Med Chem. 2013. PMID: 24059340 Review.
-
Defining the Protein Seeds of Neurodegeneration using Real-Time Quaking-Induced Conversion Assays.Biomolecules. 2020 Aug 25;10(9):1233. doi: 10.3390/biom10091233. Biomolecules. 2020. PMID: 32854212 Free PMC article. Review.
Cited by
-
Structural characterization of PHOX2B and its DNA interaction shed light on the molecular basis of the +7Ala variant pathogenicity in CCHS.Chem Sci. 2024 May 1;15(23):8858-8872. doi: 10.1039/d3sc06427a. eCollection 2024 Jun 12. Chem Sci. 2024. PMID: 38873078 Free PMC article.
-
Unfolding Mechanism and Fibril Formation Propensity of Human Prion Protein in the Presence of Molecular Crowding Agents.Int J Mol Sci. 2024 Sep 13;25(18):9916. doi: 10.3390/ijms25189916. Int J Mol Sci. 2024. PMID: 39337404 Free PMC article.