

Intracellular acidification is a hallmark of thymineless death in E. coli
- PMID: 36279294
- PMCID: PMC9632930
- DOI: 10.1371/journal.pgen.1010456
Intracellular acidification is a hallmark of thymineless death in E. coli
Abstract
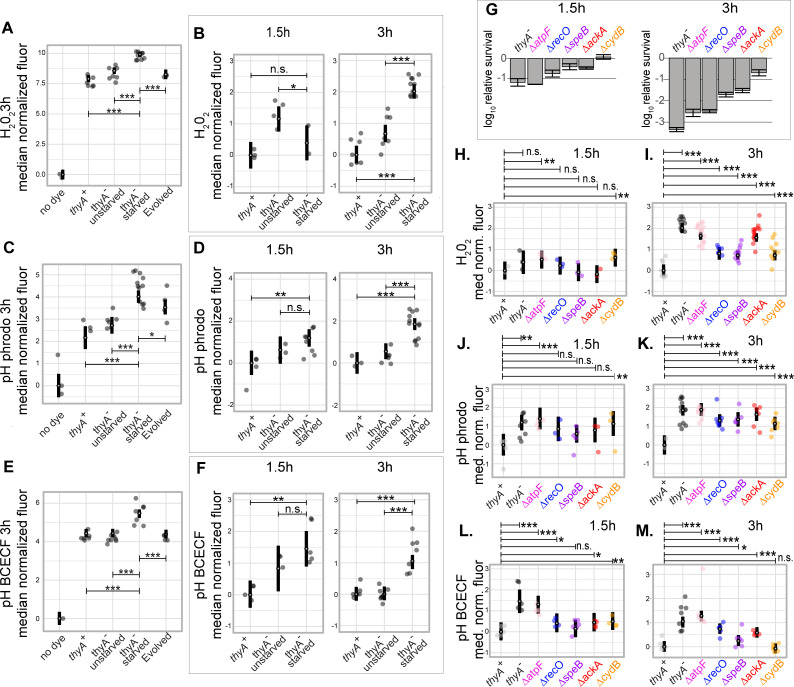
Thymidine starvation causes rapid cell death. This enigmatic process known as thymineless death (TLD) is the underlying killing mechanism of diverse antimicrobial and antineoplastic drugs. Despite decades of investigation, we still lack a mechanistic understanding of the causal sequence of events that culminate in TLD. Here, we used a diverse set of unbiased approaches to systematically determine the genetic and regulatory underpinnings of TLD in Escherichia coli. In addition to discovering novel genes in previously implicated pathways, our studies revealed a critical and previously unknown role for intracellular acidification in TLD. We observed that a decrease in cytoplasmic pH is a robust early event in TLD across different genetic backgrounds. Furthermore, we show that acidification is a causal event in the death process, as chemical and genetic perturbations that increase intracellular pH substantially reduce killing. We also observe a decrease in intracellular pH in response to exposure to the antibiotic gentamicin, suggesting that intracellular acidification may be a common mechanistic step in the bactericidal effects of other antibiotics.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures



References
-
- Wrobel A, Arciszewska K, Maliszewski D, Drozdowska D. Trimethoprim and other nonclassical antifolates an excellent template for searching modifications of dihydrofolate reductase enzyme inhibitors. J Antibiot (Tokyo). 2020;73(1):5–27. Epub 2019/10/04. doi: 10.1038/s41429-019-0240-6 ; PubMed Central PMCID: PMC7102388. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
