

Activation of the integrative and conjugative element Tn916 causes growth arrest and death of host bacteria
- PMID: 36279314
- PMCID: PMC9632896
- DOI: 10.1371/journal.pgen.1010467
Activation of the integrative and conjugative element Tn916 causes growth arrest and death of host bacteria
Abstract
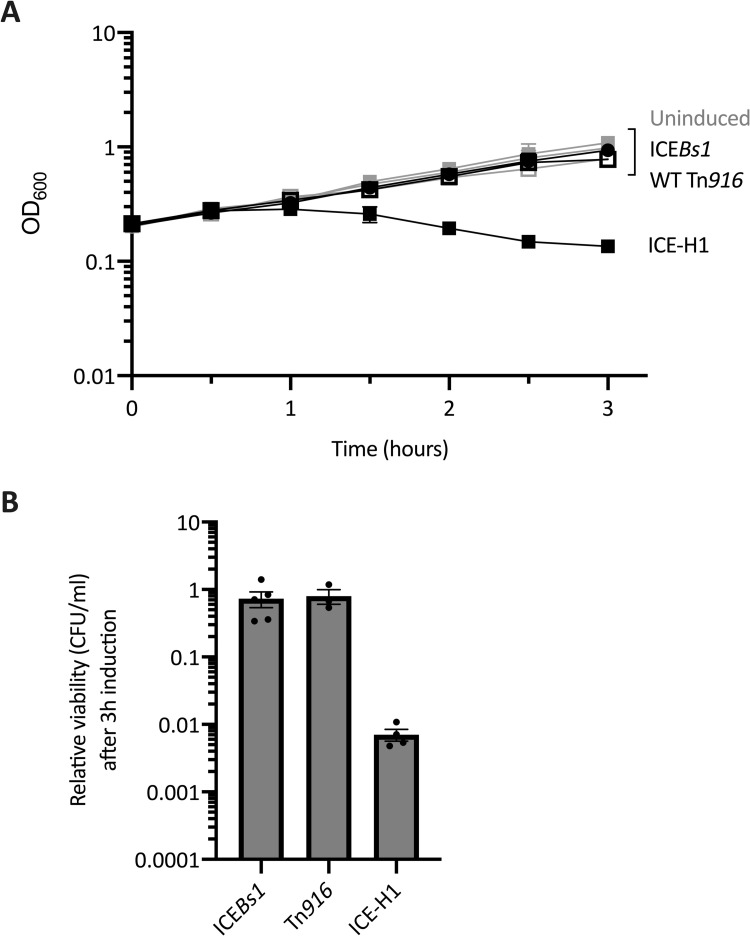
Integrative and conjugative elements (ICEs) serve as major drivers of bacterial evolution. These elements often confer some benefit to host cells, including antibiotic resistance, metabolic capabilities, or pathogenic determinants. ICEs can also have negative effects on host cells. Here, we investigated the effects of the ICE (conjugative transposon) Tn916 on host cells. Because Tn916 is active in a relatively small subpopulation of host cells, we developed a fluorescent reporter system for monitoring activation of Tn916 in single cells. Using this reporter, we found that cell division was arrested in cells of Bacillus subtilis and Enterococcus faecalis (a natural host for Tn916) that contained an activated (excised) Tn916. Furthermore, most of the cells with the activated Tn916 subsequently died. We also observed these phenotypes on the population level in B. subtilis utilizing a modified version of Tn916 that can be activated in the majority of cells. We identified two genes (orf17 and orf16) in Tn916 that were sufficient to cause growth defects in B. subtilis and identified a single gene, yqaR, that is in a defective phage (skin) in the B. subtilis chromosome that was required for this phenotype. These three genes were only partially responsible for the growth defect caused by Tn916, indicating that Tn916 possesses multiple mechanisms to affect growth and viability of host cells. These results highlight the complex relationships that conjugative elements have with their host cells and the interplay between mobile genetic elements.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
